
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Пирамидная система. Функции пирамидных систем.
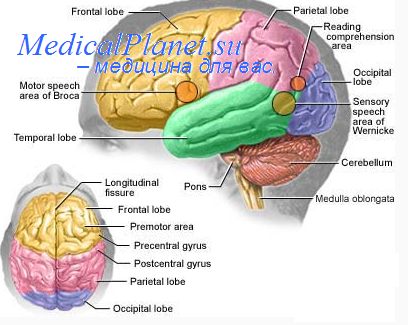
Пирамидный тракт образуют кортикоспинальные волокна, отходящие от нейронов всех моторных зон коры и направляющиеся в передние рога спинного мозга. Через каждую пирамиду продолговатого мозга проходит примерно 1 миллион кортикоспинальных волокон.
Но лишь 30—40% из них составляют волокна, отходящие от пирамидных клеток Беца в ПМК. Источником еще примерно 30% волокон являются премоторные зоны коры (ПреМК, ДМ К). Оставшиеся 30—40% волокон представляют собой отростки нейронов теменной коры, прежде всего первичной соматосенсорной коры, которые подходят к вставочным нейронам задних рогов и контролируют восходящую сенсорную афферентацию и состояние спинальных рефлексов.
Большинство в пирамидном тракте составляют тонкие волокна (диаметром 1—4 мкм). Доля крупных волокон (диаметром 10 мкм), отходящих от клеток Беца первичной моторной коры — всего лишь около 3%.
Кортикоспинальные волокна, как правило, оказывают возбуждающее влияние на спинальные нейроны, выделяя на своих окончаниях глутамат, но при этом они могут облегчать как тормозные, так и стимулирующие механизмы в сегментарном аппарате спинного мозга. Большая часть этих волокон образует синапсы непосредственно со спинальными мотонейронами передних рогов, чаще всего с той их частью, которая сосредоточена в дорсолатеральных отделах передних рогов и иннервирует дистальные отделы конечностей.
Волокна, регулирующие функцию проксимальных и аксиальных отделов, проецируются на вентромедиальную часть передних рогов и чаще контактируют со вставочными или проприоспинальными нейронами, влияя на активность мотонейронов опосредованно. Благодаря этим непрямым связям возможны более массивные, но менее дифференцированные движения, одновременно вовлекающие несколько суставов (например, вытягивание руки или ходьба).
Перекрещенные кортикоспинальные волокна, следующие в боковых столбах, преимущественно подходят к нейронам, иннервирующим мышцы дистальных отделов конечностей, тогда как неперекрещенные волокна в передних столбах чаще контактируют с вставочными нейронами обеих половин спинного мозга, иннервирующими мышцы проксимальных и аксиальных отделов.
Сравнительные исследования показывают, что число волокон в пирамидных трактах, а также доля кортикоспинальных волокон, имеющих прямые контакты с мотонейронами, у человека значительно выше, чем у приматов. Это связано с повышением сложности тонких дифференцированных действий, требующих независимых движений соседних пальцев (например, при захвате кистью небольших предметов). Но способность к таким движениям определяется не избирательностью связей корковых и спинальных мотонейронов.
Большинство кортикоспинальных нейронов связано моносинаптическими связями более чем с одним спинальным нейроном, хотя моторное поле данного коркового нейрона все же ограничено относительно небольшим количеством функционально связанных мышц (обычно 2—3). При этом некоторые кортикоспинальные нейроны одновременно оказывают разнонаправленное действие на мышцы — антагонисты.
И наоборот, передние рога данного сегмента спинного мозга получают афферентацию от различных корковых зон, хотя при этом сохраняют соматотопическую организацию. На каждом спинальном мотонейроне имеется несколько тысяч синапсов, при этом каждый корковый нейрон на данной спинальной клетке образует лишь один синапс. Чтобы инициировать активность данной двигательной единицы, часто необходима стимуляция со стороны нескольких кортикоспинальных нейронов.
Корковые нейроны, совместно влияющие на одну и ту же мышцу, часто расположены в пределах одной корковой колонки, хотя их индивидуальные моторные поля могут существенно различаться.
Как же объяснить соматотопическую специфичность и способность к тонким дифференцированным движениям, учитывая широкую конвергенцию и дивергенцию нисходящих кортикоспинальных проекций от различных корковых моторных полей? Полагают, что соматотопия на макроанатомическом уровне внутри моторной системы генетически детерминирована, но тонкая настройка нейронной сети зависит от функционального состояния синапсов, которое определяется их активностью.
Влияние на данную двигательную единицу со стороны того или иного кортикоспинального нейрона может быть дифференциально усилено или ослаблено благодаря синаптической реорганизации. Это приводит к координации функционально связанных, но пространственно удаленных корковых нейронов или их групп и может объяснять дифференцированность движения несмотря на отсутствие однозначного соответствия между корковым и спинальным мотонейроном. Условием выполнения тонких движений являются также особые свойства кортикоспинальных синапсов: при повторяющейся корковой стимуляции происходит увеличение амплитуды возбуждающих постсинаптических потенциалов в спинальных мотонейронах.
Стимуляция кортикоспинальных волокон приводит к параллельной активации альфа- и гамма-мотонейронов и, соответственно, одновременному сокращению экстра- и интрафузальных волокон в одной и той же мышце, что повышает чувствительность мышечных веретен к изменению длины мышцы (даже в условиях, когда мышца сокращена).
Селективное поражение кортикоспинальных путей (на уровне пирамид или ножек мозга), следующих от ПМК, вызывает вялый паралич конечностей, более выраженный в руках, чем в ногах. Однако затем происходит относительно быстрое восстановление функции мышц туловища и проксимальных отделов конечностей, позднее — функции дистальных отделов конечностей, однако тонкие движения в дистальных отделах конечностей, как правило, остаются нарушенными.
Таким образом, произвольные движения могут осуществляться и без участия кортикоспинального тракта, но в этом случае они утрачивают быстроту, ловкость, точность. Мышечная гипотония постепенно регрессирует, но спастичность и усиление сухожильных рефлексов после избирательного поражения кортикоспинальных волокон от ПМК не развиваются, если не повреждены следующие в составе пирамидного тракта или вблизи него (на уровне внутренней капсулы, ствола или спинного мозга) волокна от ПреМК и, возможно, ДМК, которые влияют на спинной мозг опосредованно — через ретикулоспи-нальный тракт (кортико-ретикуло-спинальные волокна).
Совместным поражением кортикоспинальных и кортико-ретикуло-спинальных волокон во многом объясняются особенности классической картины спастического паралича.
- Читать далее "Роль стволово-спинальных систем в регуляции движений. Медиальная система."
Оглавление темы "Пирамидная система. Мозжечок.":1. Поясная моторная кора. Корково-подкорковые круги.
2. Моторный круг. Сенсомоторный круг. Окуломоторный круг.
3. Взаимодействие между различными кругами головного мозга. Функции моторного круга.
4. Функции базальных ганглиев. Признаки поражения базальных ганглиев.
5. Пирамидная система. Функции пирамидных систем.
6. Роль стволово-спинальных систем в регуляции движений. Медиальная система.
7. Интерстициоспинальный тракт. Латеральная система.
8. Мозжечок. Мозжечковая система в регуляции движений.
9. Корково-мозжечковый круг. Стволово-мозжечковый круг.
10. Роль мозжечка в системе регуляции движений. Функции мозжечка.