
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Классификация цитокинов кожи
Первые описанные цитокины (например, ИЛ-1, ИЛ-2 и интерфероны) характеризовались четким и легко идентифицируемым биологическим действием. Термин «цитокины» был впервые введен Cohen в 1975 году для описания активных соединений, которые выделялись в надосадочную жидкость, получавшуюся в результате центрифугирования клеток эпителиальной линии.
До того образование подобных соединений считалось характерным только для лимфоцитов (лимфокины) и моноциов (монокины), и эти вещества считались производными иммунной системы. Цитокины кератиноцитов были впервые описаны в 1981 году, а теперь установлено, что эти клетки по числу производимых цитокинов могут соперничать с практически любым другим типом клеток в человеческом организме.
Список молекул, которые с полным правом можно назвать цитокинами, продолжает расширятся, что позволило назвать цитокинами широкий спектр биологически активных молекул, оказывающих различное действие. Развитие методов геномики позволило идентифицировать новые гены цитокинов, основываясь на их гомологии с уже известными генами этих веществ. Понять значение всего множества таких медиаторов становится все сложнее, поэтому необходима разработка стратегий, направленных на упрощение анализа вселенной цитокинов.
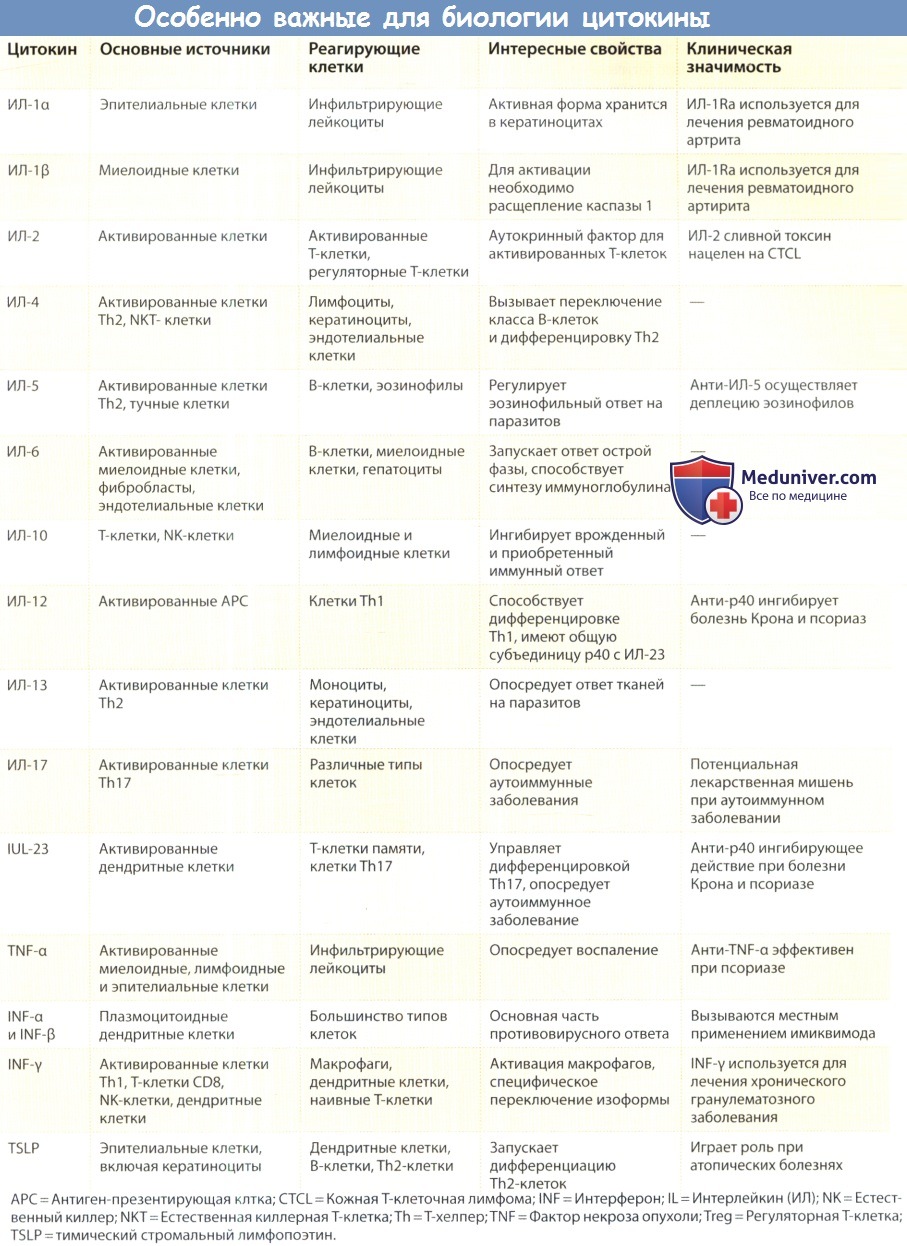
а) Первичные и вторичные цитокины. Простой концепцией, очень полезной при обсуждении функции цитокинов, продолжает оставаться концепция «первичных» и «вторичных» цитокинов. Первичными цитокинами считаются цитокины способные самостоятельно инициировать все процессы, необходимые для лейкоцитарной инфильтрации тканей. ИЛ-1 (обе формы а и β) и фактор некроза опухоли (ФИО, включая как ФНО-а, так и ФНО-β) функционируют как первичные цитокины, как и некоторые другие цитокины, которые посылают сигнал посредством рецепторов, запускающих путь нуклеарного фактора кВ (NF-кВ).
ИЛ-1 и ФИО способны вызывать экспрессию молекул клеточной адгезии на эндотелиальных клетках (селектинов, а также членов суперсемейства иммуноглобулинов, таких как молекула внутриклеточной адгезии 1 (ICAM-1) и молекула адгезии клеток сосудов (VAMC-1)], чтобы стимулировать различные клетки к производству ряда дополнительных цитокинов и вызвать экспрессию хе-мокинов, обеспечивающих хемотаксический градиент для прямой миграции специфических популяций лейкоцитов к месту воспаления (см. главу 12). Первичные цитокины можно рассматривать как часть системы врожденного иммунитета; фактически они делят сигнальные пути с толл-подобными рецепторами (TLR), семейством рецепторов, которые распознают молекулярные структуры, характерно ассоциирующиеся с микробными продуктами.
Хотя иногда и другие цитокины обладают сильной воспалительной активностью, они не дублируют эту последовательность действий целиком. Многие из них квалифицируются как вторичные цитокины, синтез которых запускается после стимуляцией ИЛ-1 и/или молекулами семейства ФИО. Термин «вторичные» не означает, однако, что такие цитокины менее важны или менее активны, чем первичные цитокины. Он скорее указывает на то, что спектр их действия имеет более ограниченный характер.
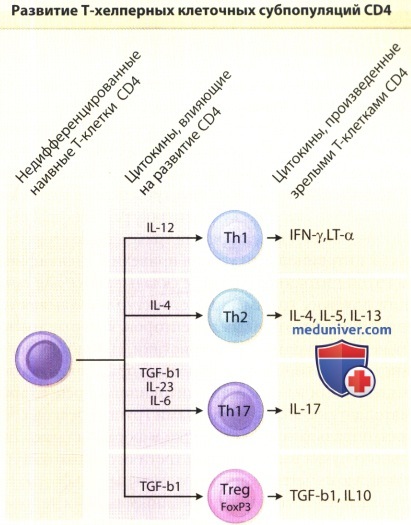
Цитокиновая среда в момент активации наивных недифференцированных Т-клеток CD4 оказывает значительное влияние на конечную модель секреции цитокинов полностью дифференцированными Т-клетками.
Субпопуляции эффекторных клеток CD4 (регуляторных Т-клеток) экспрессируют фактор транскрипции FoxP3, а их действие опосредуется частичной выработкой трансформирующего фактора роста-и1 (TGF-b1) и/или интерлейкина-10 (ИЛ-10);
INF = Интерферон; LT=Лимфотоксин.
б) Т-клеточные субпопуляции, отличающиеся по способу выработки цитокинов. Еще одной ценной концепцией, которая выдержала испытание временем, является разделение многих производимых Т-клетками цитокинов на группы в соответствии со специфическими Т-хелперными субпопуляциями, которые их вырабатывают. Две исходные Т-хелперные популяции обозначаются как Th1 и Th2. В одной из этих двух моделей секреции цитокинов участвуют также цитотоксические клетки CD8 и Т-клетки γ/δ.
Доминирование цитокинов типа 1 или типа 2 в Т-клеточном иммунном ответе имеет важные последствия для результата иммунных реакций на определенные патогены и внешние белки, способные служить аллергенами. Почти два десятилетия спустя после первого описания субпопуляций Th1 и Th2 появились убедительные данные о том, что существуют другие функционально значимые модели секреции цитокинов Т-клетками.
Наиболее важными среди этих новых популяций Т-клеток являются Тh17 и регуляторные Т-клетки (сокращенно Treg-клетки). Популяция Тh17-клеток отличается способностью синтезировать в больших количествах ИЛ-17, однако многие Тh17-клетки также секретируют ИЛ-21 и ИЛ-22. Тh17-клетки вызывают воспаление. Существуют убедительные доказательства, полученные при изучении аутоиммунных заболеваний у человека и на моделях этих болезней у мышей, согласно которым клетки, вырабатывающие ИЛ-17, являются ключевыми эффекторами при аутоиммуной патологии.
Оказалось, что популяция Т-клеток, обозначаемая как Treg, играет ключевую роль в поддержании периферической толерантности к аутоантигенам. Две из наиболее характерных черт регуляторных Т-клеток (Treg) — это экспрессия ими фактора транскрипции FохР3 и выработка трансформирующего фактора роста-β (TFG-β), цитокина, требующегося регуляторным Т-клеткам для ограничения избыточной активности провоспалительных субпопуляций Т-клеток. ИЛ-10 также вносит существенный вклад в супрессивную активность Treg-клеток, в особенности на слизистых.
Кроме того, существуют следующие типы хелперных Т-клеток: фолликулярные Т-хелперы (Tfh), которые специализируются на помощи В-клеткам в герминативных центрах; Th9-клетки, которые выделяются по признаку высокого уровня продукции ИЛ-9 и наряду с Th2 имеют противопаразитарную воспалительную активность; а также Th22-клекти, которые ассоциированы с процессом воспаления кожи, которое обусловлено именно ими, но не другими ТЫ7-ассоциированными цитокинами. Мало того, что каждый из этих подтипов Т-клеток демонстрирует свой собственный профиль продукции цитокинов, цитокины еще и являются ключевыми факторами, оказывающими влияние на дифференциацию необученных Т-клеток в описанные выше подтипы. ИЛ-12 является ключевым фактором, способствующим субпопуляции Тh1, ИЛ-4 требуется для дифференцировки Th2, а ИЛ-23 и TGF- β способствуют развитию Тh17.

в) Структурная классификация цитокинов. Не все полезные классификации цитокинов основаны исключительно на анализе их функции. Специалисты по структурной биологии, используя усовершенствованные методы синтеза гомогенных препаратов белков и новые аналитические методики (например, магнитно-резонансную спектроскопию структуры белка в растворе), дополняющие классическую методику рентгенографической кристаллографии, определи триммерную структуру многих цитокинов. Эти усилия привели к идентификации групп цитокинов, которые складываются для генерации похожих триммерных структур и связываются с группами цитокиновых рецепторов, которые имеют схожие структурные характеристики. Например, большинство цитокиновых лигандов, которые связываются с рецепторами из семейства цитокиновых рецепторов гематопоэтина, являются членами группы белков с четырехспиральным пучком.
Белки в форме четырехспирального пучка имеют общую третичную архитектуру, состоящую из четырех антипараллельных альфа-спиралей, разделенных короткими соединительными петлями.
В норме некоторые цитокины находятся в виде олигомеров, а не мономеров, что было открыто при исследовании структуры этих молекул. Например, интерферон-γ (ИНФ-γ) представляет собой цитокин в форме четырехспирального пучка, существующего естественным образом как нековалентный димер. Бивалентность димера позволяет этому лиганду связываться и олигомеризировать два комплекса рецепторов ИНФ-γ, облегчая таким образом передачу сигнала. Как ФНО-а, так и ФНО-β являются тримерами, которые состоят почти исключительно из β-листов, уложенных в структурный массив по типу «рулета с джемом».
Лигандная тримеризация рецепторов в семействе рецепторов ФНО участвует в инициации процесса сигнализации.
- Вернуться в оглавление раздела "дерматология."
Оглавление темы "Иммунология кожи.":- Регуляторные Т-клетки кожи (Treg-клетки)
- Фолликулярные Т-хелперы (Tfh-клетки) кожи
- Лимфоциты кожи в норме и при болезни
- Хоуминг Т-клеток памяти в коже
- Принципы представления антигенов антигенпредставляющими клетками кожи
- Дендритные клетки как антигенпредставляющие клетки кожи
- Клетки Лангерганса как антигенпредставляющие клетки кожи
- Дендритные клетки дермы как антигенпредставляющие клетки
- Определение и функции цитокинов кожи
- Классификация цитокинов кожи