
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Схема механизма некроза клетки
Выживание клетки зависит от поддержания постоянства ее объема и параметров внутриклеточной среды. Поскольку клеточная мембрана обладает высокой проницаемостью для воды, которая перемещается по осмотическому градиенту, поддержание объема клетки в большой мере зависит от осмотического баланса.
Для того чтобы уравновесить высокую внутриклеточную концентрацию белков, аминокислот и других органических субстратов, клетка понижает концентрацию ионов в цитозоле. Эта работа выполняется Na+/K+-АТФазой, которая выкачивает ионы Na+ из клетки в обмен на ионы К+.
В норме клеточная мембрана лишь частично проницаема для ионов Na+, но высокопроницаема для ионов К+, ввиду чего К+ вновь диффундирует наружу. Указанное истечение К+ создает внутри клетки отрицательный заряд, выталкивающий из клетки Cl-. В ходе данного перемещения ионов, сопровождающегося расходованием аденозин-5'-трифосфата (АТФ), снижение концентрации ионов Na+ и Cl- в цитозоле (приблизительно до 230 мосм/л) значительно более выражено, чем подъем концентрации К+ в цитозоле (приблизительно 140 мосм/л).
Снижение внутриклеточной концентрации Na+ при участии Na+/К+-АТФазы необходимо не только для профилактики набухания клетки, но и для создания высокого электрохимического градиента Na+, который используется для осуществления ряда транспортных процессов. Na+/К+-антипортер выводит один ион Н+ в обмен на один ион Na+, тогда как антипортер 3 Na+/Ca2+ выводит один ион Са2+ в обмен на три иона Na+.
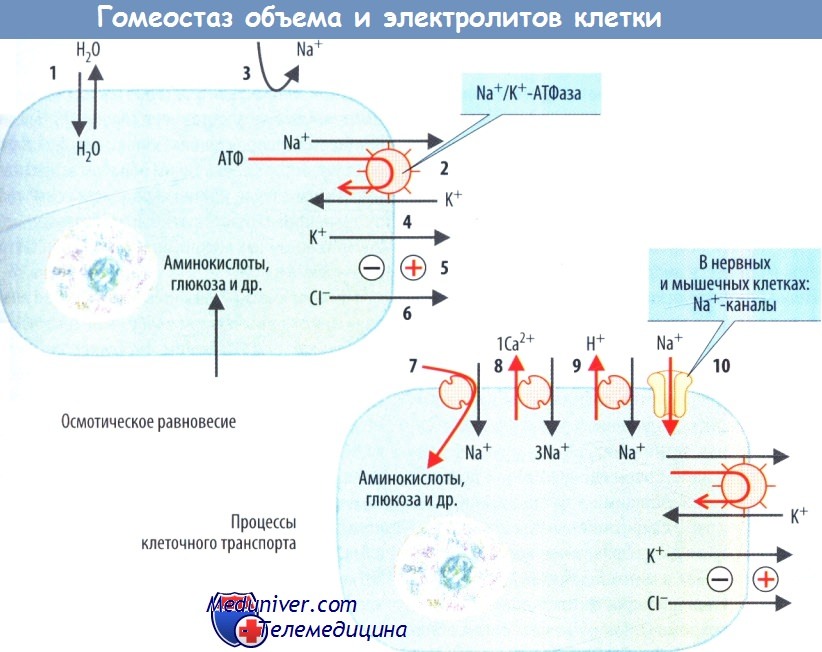
Na+-зависимые транспортные процессы также позволяют осуществлять (во вторую очередь) активное поглощение клеткой аминокислот, глюкозы, фосфатов и т.д. В конечном итоге деполяризация, достигаемая открытием Na+-каналов, обеспечивает регуляцию функции возбудимых клеток, например передачу и обработку сигнала в нервной системе или инициацию мышечных сокращений.
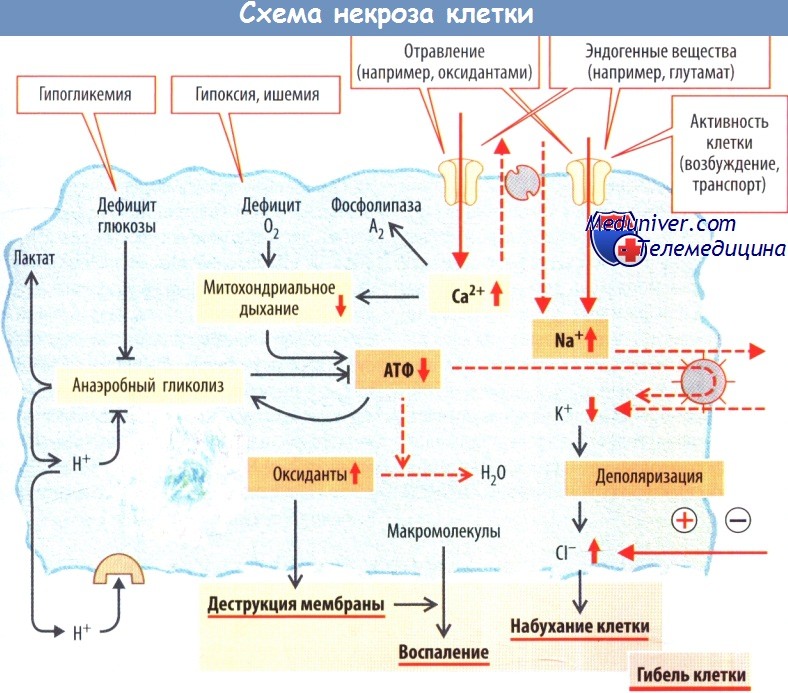
Поскольку переносчики Na+ и натриевые каналы постоянно закачивают Na+ в клетку, для выживания клетки необходима постоянная работа Na+/K+-АТФазы. Этот внутриклеточный гомеостаз Na+ может быть нарушен, если действие Na+/K+-АТФазы блокировано вследствие дефицита АТФ (при ишемии, гипоксии либо гипогликемии). В результате падает концентрация внутриклеточного К+, а уровень внеклеточного К+ растет. Клеточная мембрана деполяризуется.
Вследствие этого Cl- входит в клетку и клетка разбухает. Это происходит и в том случае, когда нарушено энергообеспечение клетки или вход Na+ превышает максимальные транспортные возможности Na+/К+-АТФазы. Многочисленные эндогенные вещества (например, нейротрансмиттер глутамат) и экзогенные яды (например, окислители) повышают вход Na+ и/или Са2+ посредством активации соответствующих каналов.
Повышение внутриклеточной концентрации Na+ не только приводит к набуханию клетки, но и к повышению концентрации Са2+ в цитозоле вследствие нарушения работы антипортера 3 Na+/Ca2+. Са2+ вызывает ряд клеточных эффектов; кроме того, он проникает в митохондрии и приводит к дефициту АТФ из-за ингибирования митохондриального дыхания.
При недостатке O2 энергетический метаболизм переключается на анаэробный гликолиз. Образование молочной кислоты, распадающейся на лактат и Н+, вызывает закисление цитозоля. Это влияет на активность внутриклеточных ферментов, в конечном итоге приводя к подавлению гликолиза, в результате исчезает последний источник АТФ. Продукция лактата приводит в дальнейшем к внеклеточному ацидозу, который влияет на функцию клетки через Н+-чувствительные рецепторы и каналы.
В условиях дефицита энергии клетка более уязвима к окислительным повреждениям, поскольку защитные механизмы клетки против окислителей (радикалов O2) АТФ-зависимы. Существует риск разрушения клеточной мембраны (перекисное окисление липидов) и высвобождения внутриклеточных макромолекул в межклеточное пространство. Поскольку иммунная система незнакома с внутриклеточными макромолекулами, к ним отсутствует иммунологическая толерантность. Иммунная система активируется, происходит дальнейшее повреждение клетки, вследствие чего развивается воспаление.
Временной промежуток, через который произойдет некротическая гибель клетки вследствие недостаточного снабжения энергией, зависит от интенсивности входа Na+ и Са2+, т. е., например, от активности возбудимых клеток или скорости транспорта эпителиальных клеток. Поскольку потенциалзависимые Na+-каналы возбудимых клеток активируются посредством деполяризации клеточной мембраны, деполяризация ускоряет гибель клетки. Гипотермия снижает активность таких каналов и тем самым замедляет процессы, приводящие к гибели клетки.
- Рекомендуем ознакомиться со следующей статьей "Схема механизма апоптоза клетки"
Оглавление темы "Патофизиология в схемах":- Физиология роста клетки и ее адаптации
- Схема внутриклеточной передачи сигнала и его нарушений
- Фосфатидилинозитол-3-киназа (PI3-киназа) зависимая передача сигнала
- Схема механизма некроза клетки
- Схема механизма апоптоза клетки
- Схема образования опухолевых клеток - онкогенеза
- Схема влияний опухоли на организм - паранеопластических синдромов
- Схема регуляции старения организма человека и длительности его жизни
- Схема повышения температуры тела - лихорадки
- Схема перегревания организма (теплового удара) и злокачественной гипертермии