
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Механизмы клеточно-опосредованной гиперчувствительности (IV типа)
Гиперчувствительность типа IV (клеточно-опосредованную гиперчувствительность) вызывают активированные антигеном (сенсибилизированные) Т-лимфоциты, включая клетки CD4+ и CD8+.
Опосредованная Т-клетками CD4+ гиперчувствительность, индуцированная антигенами окружающей среды или аутоантигенами, может быть причиной хронического воспалительного заболевания.
В настоящее время известно, что многие аутоиммунные болезни возникают в результате воспалительной реакции, вызываемой Т-клетками CD4+. При некоторых аутоиммунных болезнях, обусловленных Т-клетками, определенную роль также могут играть Т-клетки CD8+, а в случае вирусных инфекций в опосредованных Т-клетками реакциях Т-лимфоциты CD8+ могут оказаться доминирующими эффекторными клетками.
а) Реакции Т-клеток CD4+: гиперчувствительность замедленного типа и иммунное воспаление. Воспалительные реакции, вызываемые Т-клетками CD4+, первоначально были охарактеризованы как гиперчувствителъностъ замедленного типа на введенные экзогенные антигены. Те же иммунологические процессы ответственны за реакции хронического воспаления на аутоантиген.
Поскольку при такой воспалительной реакции центральную роль играет система приобретенного иммунитета, иногда эту реакцию называют иммунным воспалением. Тh1-клетки и Тh17-клетки вносят свой вклад в органоспецифические заболевания, для которых воспаление является важным элементом патологии.
При воспалительной реакции, ассоциированной с Тh1-клетками, немалую роль играют активированные макрофаги, тогда как в случае воспаления, индуцированного Тh17-клетками, доминирует нейтрофильный компонент.
Клеточные процессы при опосредованной Т-клетками гиперчувствительности включают серию реакций, в которых существенная роль принадлежит цито-кинам. Эти реакции имеют несколько стадий.
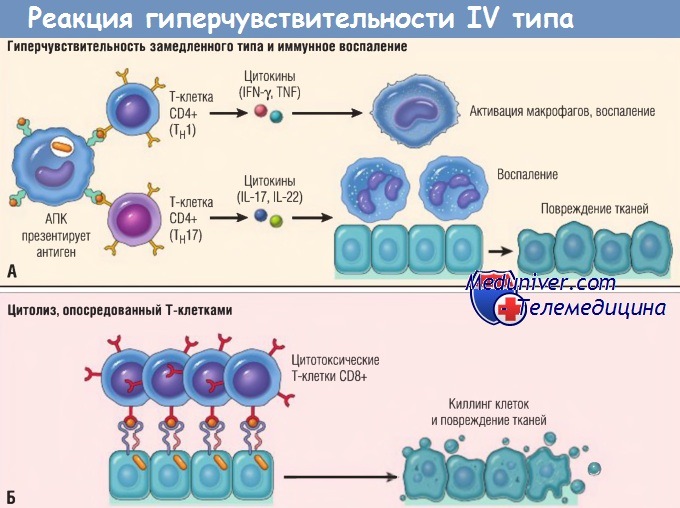
(А) В реакциях гиперчувствительности замедленного типа Тн1-клетки CD4+ (а иногда Т-клетки CD8+, не показаны) отвечают на тканевые антигены секрецией цитокинов,
которые стимулируют воспаление и активируют фагоцитоз, приводя к повреждениям тканей. Тн17-клетки CD4+ способствуют воспалению, мобилизуя нейтрофилы (в меньшей степени моноциты).
(Б) При некоторых заболеваниях цитотоксические Т-клетки CD8+ непосредственно убивают клетки тканей.
IFN — интерферон; IL — интерлейкин; TNF — фактор некроза опухоли; АПК — антигенпрезентирующая клетка.
б) Пролиферация и дифференцировка Т-клеток CD4+. Наивные Т-клетки CD4+ распознают пептиды, презентированные дендритными клетками, и секретируют IL-2, который функционирует как аутокринный фактор роста, стимулирующий пролиферацию антиген-реактивных Т-клеток. Последующую дифференцировку стимулированных антигеном Т-клеток в Тh1-клетки или Th17-клетки индуцируют цитокины, продуцируемые АПК во время активации Т-клеток.
В некоторых случаях АПК (дендритные клетки и макрофаги) образуют IL-12, индуцирующий дифференцировку Т-клеток CD4+ в субпопуляцию Тh1-клеток. IFN-y, продуцируемый этими эффекторны-ми клетками, стимулирует дальнейшее развитие Тh1-клеток, тем самым усиливая реакцию.
Если АПК образуют воспалительные цитокины, такие как IL-1, IL-6 и IL-23 (близкородственный IL-12), они все вместе с TGF-P (продуцируемым клетками многих типов) стимулируют дифференцировку Т-клеток в субпопуляцию Тh17-клеток.
Некоторые из дифференцированных эффекторных клеток поступают в кровоток и могут оставаться в пуле Т-клеток памяти в течение длительного срока, иногда исчисляемого годами.
в) Иммунный ответ дифференцированных эффекторных Т-клеток. При повторном воздействии антигена предварительно активированные Т-клетки распознают его после презентации АПК и отвечают на него. Тh1-клетки секретируют цитокины, в основном IFN-y, ответственные за многие проявления гиперчувствительности замедленного типа.
Активированные IFN-y макрофаги претерпевают ряд изменений: заметно возрастает способность макрофагов к фагоцитозу и киллингу микроорганизмов; увеличивается число молекул МНС класса II, экспрессируемых на поверхности макрофагов, и тем самым повышается эффективность дальнейшей презентации антигена; макрофаги секретируют TNF, IL-1 и хемокины, способствующие воспалению; наконец, макрофаги продуцируют дополнительное количество IL-12, усиливая ответ Тh1-клеток.
Таким образом, активированные макрофаги служат инструментом элиминации патогенного антигена; если активация продолжается, воспаление не прекращается и происходит поражение тканей.
Тh17-клетки активируются некоторыми микробными антигенами и аутоантигенами при аутоиммунных заболеваниях. Активированные Тh17-клетки секретируют IL-17, IL-22, хемокины и другие цитокины. Цитокины вовлекают в реакцию нейтрофилы и моноциты, тем самым способствуя воспалению. Тh17-клет-ки также продуцируют IL-21, усиливающий ответ.
Классическим примером гиперчувствительности замедленного типа служит туберкулиновая реакция, возникающая при внутрикожном введении очищенного деривата белка (называемого также туберкулином), содержащего белок антигена туберкулезных микобактерий. У предварительно сенсибилизированных индивидов через 8-12 час в месте введения возникают покраснение и уплотнение тканей, достигающие максимума спустя 24-72 час, после чего эти проявления медленно исчезают.
Морфологически гиперчувствительность замедленного типа характеризуется накоплением мононуклеарных клеток, в основном Т-клеток CD4+ и макрофагов, образующих вокруг венул периваскулярные скопления. При полностью развившейся реакции обнаруживают заметную гипертрофию эндотелия, отражающую опосредованную цитокинами активацию эндотелиальных клеток.
В случае антигенов, которые персистируют длительное время и не подвергаются разрушению (например, туберкулезных микобактерий, колонизирующих легкие или другие ткани), в периваскулярном инфильтрате в течение 2-3 нед доминируют макрофаги. Активированные макрофаги часто претерпевают морфологическую трансформацию, превращаясь во внешне сходные с эпителием клетки — эпителиоидные клетки.
Микроскопическую агрегацию эпителиоидных клеток, обычно окруженных кольцом, состоящим из лимфоцитов, называют гранулемой. Эта форма воспаления, называемая гранулематозным воспалением, обычно ассоциирована с интенсивной активацией Т-клеток, сопровождающейся продукцией цитокинов. Такое воспаление также может быть вызвано инородными телами, активирующими макрофаги, в отсутствие приобретенного иммунного ответа.
Контактный дерматит представляет собой типичный пример повреждения тканей, вызываемого реакцией гиперчувствительности замедленного типа. Он может возникнуть вследствие контакта с урушиолом, антигенным компонентом яда сумаха укореняющегося, и проявляется везикулярным дерматитом.
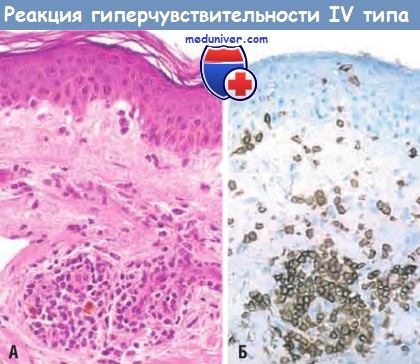
(А) Периваскулярная инфильтрация Т-клетками и мононуклеарными фагоцитами.
(Б) Иммунопероксидазная реакция выявляет преимущественно периваскулярный клеточный инфильтрат, положительно реагирующий с антителами, специфическими к CD4.
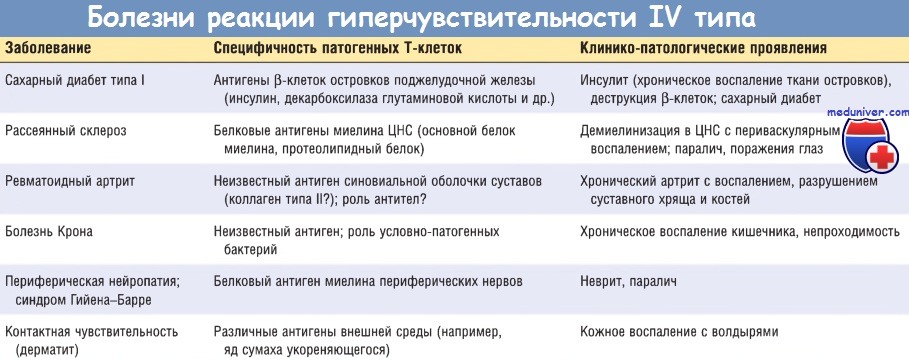
г) Реакции Т-клеток CD8+: клеточно-опосредованная цитотоксичность. В этой реакции, опосредованной Т-клетками, цитотоксические Т-лимфоциты CD8+ уничтожают несущие антиген клетки-мишени. Разрушаемые под влиянием цитотоксических Т-лимфоцитов ткани могут быть важным компонентом многих опосредованных Т-клетками заболеваний, в частности сахарного диабета типа I.
Цитотоксические Т-лимфоциты, специфические для антигенов гистосовместимости, присутствующих на клеточной поверхности, играют немаловажную роль в отторжении трансплантатов. Цитотоксические Т-лимфоциты также принимают участие в реакциях против вирусов. В клетках, инфицированных вирусами, молекулы МНС класса I презентируют вирусные пептиды и комплекс пептид-МНС класса I, а также распознают TCR Т-лимфоцитов CD8+.
Киллинг инфицированных клеток устраняет инфекцию, но приводит к повреждению клеток (например, при вирусном гепатите). На поверхности клеток также презентированы опухоль-ассоциированные антигены, и цитотоксические Т-лимфоциты участвуют в отторжении опухолей.
Ведущий механизм опосредованного Т-клетками киллинга клеток-мишеней связан с действием перфоринов и гранзимов (преформированных медиаторов, содержащихся в подобных лизосомам гранулах цитотоксических Т-лимфоцитов). Цитотоксические Т-лимфоциты, распознавшие клетки-мишени, секретируют комплекс, состоящий из перфорина, гранзимов и белка, называемого серглицином. Комплекс попадает в клетки-мишени в результате эндоцитоза. В цитоплазме клеток-мишеней перфорин стимулирует высвобождение гранзимов из комплекса.
Гранзимы представляют собой протеазы, которые расщепляют и активируют каспазы, индуцирующие апоптоз клеток-мишеней. Активированные цитотоксические Т-лимфоциты также экспрессируют лиганд Fas — молекулу, гомологичную TNF, которая связывается с Fas, экспрессированным клетками-мишенями, и индуцирует апоптоз.
Т-клетки CD8+ также продуцируют цитокины, в частности IFN-y, и участвуют в воспалительных реакциях, напоминающих реакцию гиперчувствительности замедленного типа, особенно после вирусных инфекций и воздействия некоторых контактных сенсибилизирующих агентов.
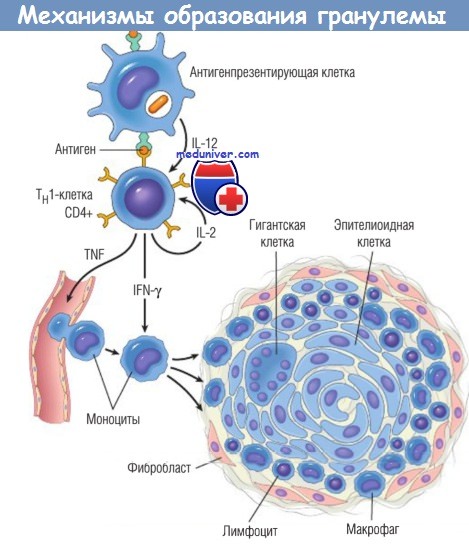
Схематическое изображение процессов, ведущих к формированию гранулем при клеточноопосредованных реакциях гиперчувствительности типа IV.
Обратите внимание на роль цитокинов.
IFN — интерферон; IL — интерлейкин; Тh — хелперные Т-клетки; TNF — фактор некроза опухоли.
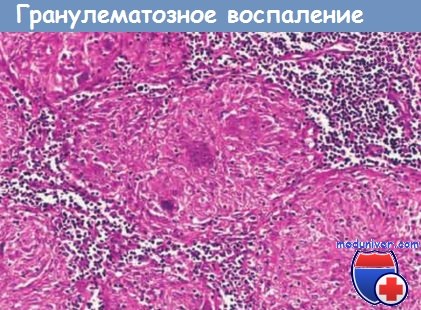
Срез лимфоузла, на котором видно несколько гранулем; каждая из них состоит из агрегата эпителиоидных клеток, окруженного лимфоцитами.
Гранулема в центре содержит несколько многоядерных гигантских клеток.
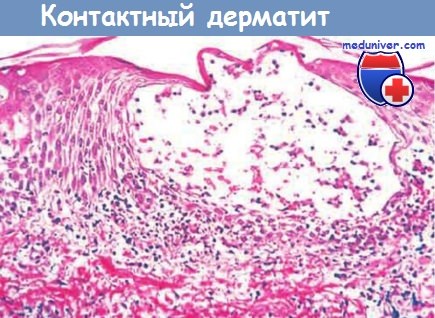
Поражение представляет собой волдырь (везикулу) с дермальным и эпидермальным мононуклеарным инфильтратом.
Видео схема реакции гиперчувствительности IV типа (аллергической реакции четвертого типа)
- Рекомендуем ознакомиться со следующей статьей "Виды аутоиммунных болезней"
Оглавление темы "Аутоиммунные болезни":- Механизмы клеточно-опосредованной гиперчувствительности (IV типа)
- Виды аутоиммунных болезней
- Иммунологическая толерантность и ее механизмы
- Патогенез (механизм) аутоиммунитета
- Эпидемиология и критерии системной красной волчанки (СКВ)
- Причины и механизмы развития системной красной волчанки (СКВ)
- Морфология системной красной волчанки (СКВ)
- Клиника системной красной волчанки (СКВ)
- Причины и механизмы лекарственной красной волчанки
- Причины и механизмы развития синдрома Шегрена