
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Строение, функции и сокращение гладкой мускулатуры - с точки зрения физиологии человека
Оглавление статьи:- Функции гладкой мускулатуры
- Структура гладкой мускулатуры
- Процесс сокращения
- Коротко
- Список использованной литературы
а) Функции гладкой мускулатуры. Гладкомышечные клетки являются составной частью стенок внутренних органов, за исключением сердца; их механические свойства приспособлены к функциям тех или иных органов.
1. Гистологические особенности. Эти мышцы называются гладкими, потому что при наблюдении в световой микроскоп в них не выявляется поперечная исчерченность. Гладкие мышцы состоят из веретеновидных клеток длиной 50-400 мкм и диаметром 2-10 мкм, с центральным ядром (рис. ниже).
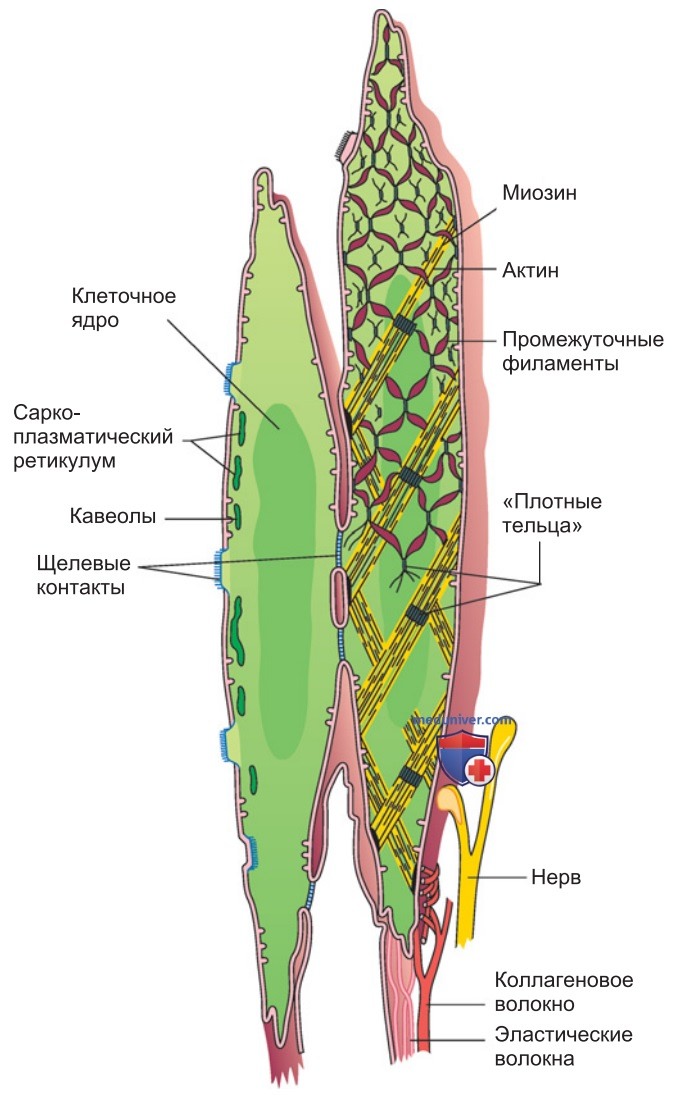
Индивидуальные мышечные клетки механически соединены особыми клеточными контактами (десмосомами), а также связаны в сеть эластическими и коллагеновыми волокнами с внеклеточным матриксом.
2. Функции, специфичные для органов. Особенности сокращения гладкой мускулатуры обусловлены функциями соответствующего органа. Здесь следует отметить перистальтику желудочно-кишечного тракта, схватки миометрия во время родов, обусловленные фазово-ритмическими сокращениями, или длительные тонические сокращения кровеносных сосудов.
Тонические сокращения внутренних сфинктеров предотвращают нежелательный выход содержимого мочевого пузыря и кишечника. Эти примеры демонстрируют функциональную роль гладкой мускулатуры.
Кроме того, становится понятным, почему гладкую мускулатуру сложно представить в качестве самостоятельного образования. Далее будут рассмотрены основные принципы ее деятельности и регуляции. Особенности, свойственные конкретным органам, следует искать в соответствующих главах (см. ссылки).
б) Структура гладкой мускулатуры. Гладкая мускулатура подразделяется структурно и функционально на два типа: унитарный (singleunit, монофункциональный однородный тип) и мультиунитарный (multi-unit, полифункциональный неоднородный тип). Гладкомышечные клетки содержат сеть из филаментов актина и миозина; степень выраженности СР разнообразна.
1. Унитарный тип. Гладкомышечные клетки этого типа электрически связаны друг с другом низкоомными контактами — нексусами, или щелевыми контактами. Таким образом, создается функционально единое образование, подобное функциональному синцитию миокарда. К этому подтипу принадлежат, среди прочих, мышцы кишечника, матки, мочеточника и некоторых кровеносных сосудов.
Они обладают спонтанной фазово-ритмической активностью миогенной природы, которую генерируют пейсмекерные клетки и модулируют вегетативные нервы.
2. Мультиунитарный тип. В мультиунитарной гладкомышечной ткани каждая клетка сокращается независимо от другой. К ней относятся мышцы радужной оболочки и ресничного (цилиарного) тела глаза, семенного протока, а также пиломоторные (волосковые) мышцы. Иногда единое функциональное образование представляет собой небольшую группу мышечных клеток, соединенных нексусами (щелевыми контактами; см. выше).
Гладкие мышцы или их группы снабжены ветвями вегетативных (автономных) нервов, в той или иной мере контактирующими с гладкомышечными клетками. Из варикозов по ходу вегетативных аксонов высвобождаются возбуждающие или тормозные нейромедиаторы. Плотность иннервации и количество щелевых контактов бывают различными. В мультиунитарных гладких мышцах спонтанная активность либо вовсе отсутствует, либо она очень слабая, так что тонус гладких мышц этого типа является преимущественно нейрогенным.
3. Смешанные подтипы. Для многих гладких мышц (например, мышц сосудов) не удается четко разграничить унитарный и мультиунитарный типы, поскольку нейрогенный мышечный тонус может проявляться в сочетании со спонтанным миогенным тонусом (базовым тонусом).
4. Ультраструктура гладкомышечных клеток. Наряду с актиновыми и миозиновыми филаментами в гладкомышечных клетках присутствуют филаменты промежуточного типа диаметром 10 нм (рис. выше). Актиновые филаменты прикреплены либо к плазматической мембране, либо к многочисленным внутриклеточным плотным тельцам (dense bodies), функциональным аналогам Z-пластинок поперечно-полосатых мышц. Актиновые филаменты вместе с миозиновыми образуют минисаркомеры. Промежуточные филаменты тоже связаны с эластичной сетью цитоскелета.
Элементы цитоскелета ориентированы диагонально по отношению к длинной оси мышечной клетки (рис. выше).
Саркоплазматический ретикулум гладкой мускулатуры, аккумулирующий Са2+, представляет собой сеть трубочек, которая по сравнению с СР скелетной мышцы менее развита и не имеет специфической организации, коррелирующей с локализацией актиновых и миозиновых филаментов; однако эта сеть местами может иметь такой же объем, как СР скелетных мышц. СР частично располагается под цитолеммой, частично в глубине клетки. Многие участки СР, находящиеся около цитолеммы, примыкают к кавеолам — углублениям клеточной мембраны (рис. выше).
Эти углубления, аналогично Т-трубочкам в поперечно-полосатых мышцах, сообщаются с межклеточным пространством, однако типичная Т-система отсутствует. Поблизости от кавеол особенно плотно распределены белки-переносчики Са2+, а также рецепторы, при активации которых высвобождается Са2+ из СР. Это позволяет предполагать, что кавеолы играют важную роль в процессе электромеханического сопряжения.
в) Процесс сокращения. Гладкие мышцы, так же как скелетные мышцы, сокращаются в результате скольжения филаментов, однако это происходит гораздо медленнее и не требует столь значительных затрат энергии.
1. Механизм сокращения. Так же как скелетная мускулатура, гладкомышечные клетки сокращаются посредством продвижения филаментов актина и миозина относительно друг друга с помощью циклической работы поперечных мостиков миозина. В то же время гидролиз АТФ под влиянием АТФазы миозина и продвижение филаментов осуществляются в 100-1000 раз медленнее, чем в быстрой скелетной мышце. Соответственно меньше затраты АТФ и потребление кислорода.
При одном и том же диаметре мышцы гладкая мускулатура может развивать более значительную силу и дольше поддерживать ее, чем скелетная мускулатура, затрачивая при этом в 100-500 раз меньше энергии.
Отсюда следует, что гладкие мышцы приспособлены для длительного поддержания тонуса, не сопровождаемого утомлением и значительными затратами энергии. Это соответствует характеру функционирования мышц крупных артерий, которые год за годом должны поддерживать кровяное давление. Такие медленные гладкие мышцы получили название тонических в противоположность фазическим гладким мышцам, например мышцам желудочно-кишечного тракта и мочеполовой системы, для которых свойственна ритмическая активность.
Однако следует учитывать, что между тонической и фазной активностью, в их чистом виде, существует множество переходных форм.
2. Мышечные белки. Небольшая скорость сокращения и экономичные затраты АТФ обусловлены чрезвычайно низкой активностью АТФазы и низким содержании миозина в гладких мышцах. Они содержат в 5 раз меньше миозина, но больше актина, чем поперечно-полосатые мышцы. В актиновых филаментах есть тропомиозин, но стоит отметить, что отсутствует тропонин. Вместо этого Са2+-белка выявлен кальмодулин, который действует в гладкой мускулатуре как сенсор Са2+.
P.S. Кальдесмон и кальпонин. В актиновых филаментах можно также обнаружить кальдесмон и кальпонин. Возможно, эти белки участвуют в процессе сокращения в качестве регуляторов. В условиях in vitro они обычно подавляют активность АТФазы актомиозина. Этот тормозный эффект усиливают ионы Са2+.
г) Коротко. Гладкая мускулатура. Гладкие мышцы лишены типичной для скелетной и сердечной мускулатуры поперечной исчерченности, поскольку актиновые и миозиновые филаменты располагаются нерегулярно. Структурно и функционально гладкие мышцы подразделяются на два основных типа. Мышцам унитарного типа свойственны фазно-ритмические сокращения миогенной природы и электрическая связь между мышечными клетками посредством низкоомных контактов, или нексусов (щелевых контактов).
Мультиунитарный тип характеризуется нейрогенным мышечным тонусом, когда каждая гладкомышечная клетка сокращается независимо от другой. Для тонических гладких мышц кровеносных сосудов невозможно установить границу между унитарным и мультиунитарным типами. При сокращении гладких мышц актиновые и миозиновые филаменты продвигаются («скользят») относительно друг друга, однако этот процесс, как и расщепление АТФ, происходит намного медленнее, чем в скелетной мускулатуре, и, следовательно, требует гораздо меньшей затраты энергии.
Таким образом, гладкие мышцы приспособлены для поддержания активности, исключающей утомление. Вместо тропонина С в качестве сенсора Са2+ выступает кальмодулин.
Видео лекция гистология мышечной ткани (гладкой, поперечнополосатой, сердечной)
- Рекомендуем ознакомиться далее "Регуляция сокращений гладкой мускулатуры - с точки зрения физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 17.10.2024