
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Энергетика сокращения скелетной мышцы - с точки зрения физиологии человека
Оглавление статьи:- Источник энергии для мышечной деятельности
- Активность АТФазы и типы мышечных волокон
- Мышечная теплота и энергетический обмен
- Коротко
- Список использованной литературы
а) Источник энергии для мышечной деятельности. В рабочем цикле поперечных мостиков АТФ служит непосредственным источником энергии; запас АТФ в мышце пополняется за счет трех различных механизмов: прямого фосфорилирования, гликолиза, окислительного фосфорилирования.
1. Возобновление АТФ. Аденозинтрифосфат расщепляется в мышце АТФазой миозина до АДФ и фосфата. Небольшой запас АТФ, имеющийся в мышечной клетке к началу активности, достаточен лишь для нескольких сокращений. Однако в мышце действуют три различных механизма пополнения запаса АТФ (табл. 2):
- прямое фосфорилирование АДФ фосфатной группой креатинфосфата;
- анаэробное образование АТФ посредством гликолиза (2-3 моль АТФ на 1 моль глюкозы);
- аэробная регенерация АТФ путем окислительного фосфорилирования (~30 моль АТФ на 1 моль глюкозы) в митохондриях.
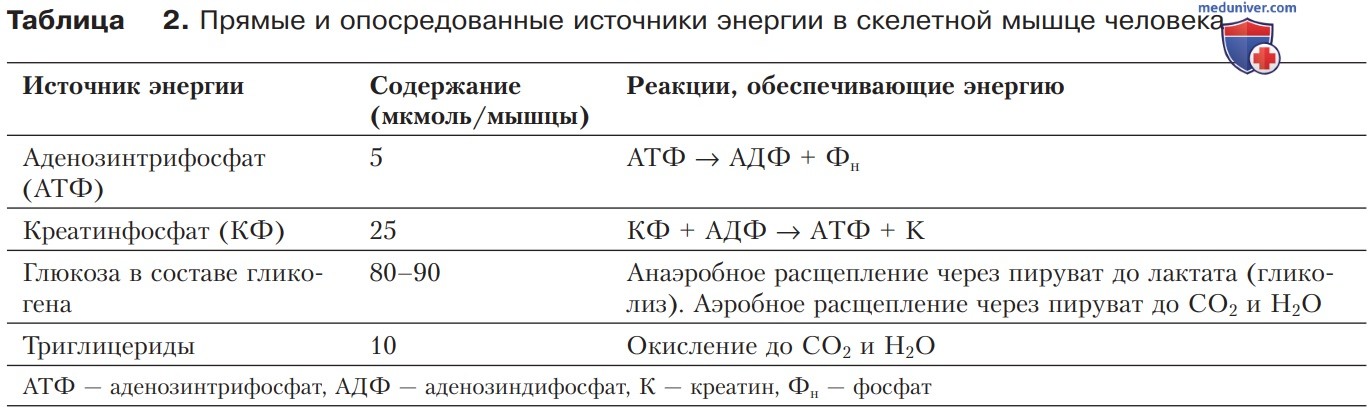
2. Ресинтез АТФ за счет креатинфосфата. Чрезвычайно быстрый процесс ресинтеза АТФ (в течение 10-20 с) путем переноса фосфатной группы от креатинфосфата к АДФ (реакция Ломана) служит своего рода буфером, обеспечивающим необходимое внутриклеточное содержание АТФ в самом начале сократительной активности.
3. Гликолиз. При интенсивной и продолжительной механической работе требуется ресинтез АТФ. В течение короткого времени (несколько минут) ресинтез АТФ может быть обеспечен с относительно высокой скоростью благодаря расщеплению глюкозы, входящей в состав гликогена (табл. 2). Однако энергетические ресурсы, освобождаемые при анаэробном гликолизе, ограничены; они достигают предела в течение ~30 с.
Гликолиз приводит к накоплению молочной кислоты в межклеточной жидкости и крови. В итоге развивается метаболический ацидоз, который ограничивает работоспособность и вызывает утомление.
4. Аэробный энергетический обмен. В самом начале продолжительной мышечной активности осуществляется аэробное образование АТФ (с некоторой задержкой — через 30-60 с после начала мышечной работы). Это происходит в результате окислительного фосфорилирования при повышенном потреблении О2 (дыхательная цепь). Энергия, необходимая для ресинтеза АТФ, поступает от окисления углеводов или жиров (табл. 2).
Если при длительной работе скорости образования и расщепления АТФ сравниваются, наступает стационарное состояние системы (steady state), когда уровни АТФ и креатинфосфата в цитоплазме постоянны. При интенсивных физических упражнениях (например, у спортсменов) скорость расщепления АТФ может увеличиваться в 100 раз. Чтобы во время продолжительной работы поддерживалась постоянная мощность (т. е. стационарное состояние), должна также увеличиваться скорость ресинтеза АТФ посредством окислительного фосфорилирования.
Поскольку для ресинтеза АТФ требуется кислород (~1/6 моль О2 на 1 моль АТФ), потребление О2 возрастает так, что может в 100 раз превысить уровень покоя. Соответственно в работающих мышцах увеличивается скорость расщепления жирных кислот и гликогена.
Аэробный синтез поставляет гораздо более значительное количество АТФ в расчете на 1 моль глюкозы, но в 2-3 раза медленнее, чем при гликолизе. В аэробных условиях расщепление АТФ и механическая эффективность мышцы в 2-3 раза ниже, чем при гликолизе. Поэтому, с одной стороны, стайер при средней скорости ~5 м/с бежит на длинной дистанции едва ли не в 2 раза медленнее, чем спринтер на короткой дистанции.
С другой стороны, в результате кратковременного усилия (например, при рывке на финишной прямой) стайер способен превысить характерную для него скорость за счет гликолитического расщепления дополнительного количества гликогена. При этом усиливается как образование, так и расщепление АТФ.
5. Погашение кислородной задолженности. До тех пор пока в процессе длительной работы не начнется аэробный ресинтез, покрывающий расход АТФ, содержание креатинфосфата в цитоплазме падает вследствие его затраты в реакции Ломана. Запас креатинфосфата не пополняется до тех пор, пока не закончится сокращение мышцы и реакция Ломана не пойдет в обратном направлении. Необходимый для этого АТФ образуется в первые минуты расслабления мышцы по окончании мышечной работы в результате окислительного фосфорилирования, т.е. реакции с потреблением О2.
Поступающий в это время О2 частично обеспечивает погашение кислородной задолженности, в т. ч. возмещение дефицита О2, обусловленного анаэробным процессом гликолиза.
б) Активность АТФазы и типы мышечных волокон. От активности АТФазы миозина зависит характер сокращений мышцы; красные мышцы богаты миоглобином и укорачиваются медленно, белые мышцы содержат мало миоглобина и укорачиваются быстро, но подвержены утомлению.
1. Скорость расщепления АТФ. Скорость укорочения мышцы тем выше, чем быстрее совершаются рабочие циклы поперечных мостиков. Скорость рабочего цикла зависит от активности изоформ АТФазы миозина. Миозины быстрых мышц расщепляют большее количество АТФ в единицу времени, чем миозины медленных мышц.
2. Типы мышечных волокон. Характер сокращения мышцы в существенной мере зависит от состава изоформ миозина (прежде всего от их АТФаз-ной активности). Большинство мышц, особенно у человека, содержат несколько (два и более) типов мышечных волокон, отличающихся по изоформам миозина. Есть три основных типа мышечных волокон: медленные волокна I типа, быстрые — IIА типа и самые быстрые — IIX типа. Последние имеют альтернативное название волокон IID типа и соответствуют волокнам IIB типа, которые встречаются у мелких млекопитающих, но не у человека. Однако скорость сокращения волокон IIX несколько меньше, чем у волокон IIB типа.
Индивидуальные мышечные волокна человека могут содержать единственную изоформу миозина либо две-три различные изоформы миозина (гибридные волокна). Типы мышечных волокон различаются не только по их АТФазной активности, но и по другим функциональным, структурным, биохимическим характеристикам, например по содержанию ферментов окислительного и гликолитического метаболизма, а также по присутствию миоглобина — родственного гемоглобину белка, обеспечивающего поступление О2 в миоциты (табл. 3).

От миоглобина зависит цвет мышц: белые мышцы содержат мало миоглобина, красные — много; кроме того, есть много смешанных типов мышц.
Красные мышцы, например мускулатура торса или камбаловидная мышца, в основном состоят из медленных волокон I типа с низкой активностью АТФазы миозина (табл. 3). Соответственно они отличаются низкой утомляемостью, малыми затратами энергии, выносливостью. Быстрые мышцы, имеющие белый или розоватый цвет (например, поясничная мышца, латеральная мышца бедра), содержат преимущественно волокна IIA и IIX типов с повышенной активностью АТФазы миозина.
В то время как волокна IIA, подобно волокнам I типа, могут благодаря особенностям своего метаболизма долго удерживать нагрузку, гликолитические белые волокна IIX типа быстро утомляются и не способны к удержанию груза или длительной работе. Волокна IIX типа совершают только быстрые сильные движения, причем запас АТФ возобновляется главным образом анаэробным путем и в мышцах накапливается молочная кислота.
3. Утомление. В развитии мышечного утомления при продолжительных или частых и сильных сокращениях участвуют различные механизмы. Утомление можно определить как ослабленную способность к поддержанию заданной силы. Это объясняется уменьшением количества Са2+, высвобождаемого из саркоплазматического ретикулума, а также нарушением взаимодействия миофибрилл с Са2+ из-за накопления молочной кислоты (внутриклеточный ацидоз) и фосфата.
Связанные с утомлением изменения рН и повышение концентрации таких метаболитов, как фосфат и АДФ, можно проследить в мышечных клетках in situ с помощью метода ядерного магнитного резонанса (ЯМР-спектроскопии). Накопление метаболитов, прежде всего молочной кислоты, не признается, как считалось ранее, причиной синдрома отсроченной мышечной боли (см. 4). Такая постепенно нарастающая боль возникает из-за микроповреждений мышц, подвергаемых непривычной интенсивной нагрузке.
в) Мышечная теплота и энергетический обмен. Мышцы с высокой эффективностью преобразуют химическую энергию в механическую энергию и тепло.
1. Энергетический обмен и теплообразование. При активации мышцы усиливается расщепление АТФ; интенсивность метаболизма в мышечной ткани возрастает в 100-1000 раз. Согласно первому закону термодинамики (закон сохранения энергии), химическая энергия, преобразуемая в мышце, должна быть равна сумме механической энергии (мышечной работы) и выделяемой теплоты. Даже в отсутствие физически измеряемой внешней мышечной работы (например, при изометрическом тетанусе, когда тело неподвижно находится в положении стоя) химическая энергия в мышце преобразуется в тепло (теплоту изометрического напряжения).
Это означает, что в результате непрерывного циклического взаимодействия с актином поперечные мостики совершают в мышце значительную «внутреннюю» статическую работу. Причем более продолжительное статическое усилие сопровождается утомлением. Когда мышца поднимает груз, совершая внешнюю работу, она преобразует дополнительное количество АТФ и производит теплоту укорочения. Таким образом, дополнительное образование энергии в процессе изотонического сокращения пропорционально выполненной работе (эффект Фенна).
Следует заметить, что тепло, выделяемое скелетными мышцами, играет роль в терморегуляции; примером служит холодовая дрожь — непроизвольные ритмичные сокращения скелетных мышц при охлаждении организма.
2. Эффективность. Гидролиз одного моля АТФ обеспечивает ~60 кДж энергии. Лишь 40-50% этой энергии сократительный аппарат преобразует в механическую энергию или работу; остальная часть выделяется в виде тепла в начале и во время сокращения мышцы, температура которой при этом несколько повышается. Преобразование энергии осуществляется в миофибриллах с коэффициентом эффективности ~40-50%.
Однако механическая эффективность деятельности всей мышцы составляет не более 20-30%, поскольку вне миофибрилл во время сокращения и после него идут восстановительные внутриклеточные процессы, которые сопровождаются теплообразованием (теплота восстановления) и требуют значительных затрат энергии. Сюда относятся активность ионных насосов и окислительная регенерация АТФ. Чем больше произведенная работа, тем выше теплообразование, а также расход источников энергии и кислорода.
1. Энергетика сокращения скелетной мышцы. АТФ служит непосредственным источником энергии сокращения мышцы. Обновление запаса АТФ в мышце происходит благодаря трем различным механизмам: переносу фосфатной группы от креатинфосфата к АДФ (реакция Ломана), гликолизу, окислению жирных кислот и углеводов. При аэробном синтезе АТФ потребление О2 зависит от мышечной деятельности; во время продолжительной стационарной деятельности затраты О2 могут быть в 100 раз выше, чем в покое.
2. Особенности сокращения различных типов мышц. Характер сокращения мышцы зависит от того, из каких волокон она состоит: из волокон быстрого типа (IIA, IIX типы) или медленного типа (I тип). Продолжительную деятельность и работу по удержанию нагрузки эффективнее осуществляют медленные мышцы (обычно красные мышцы, поскольку содержат много миоглобина). Быстрые мышцы (белые мышцы с низким содержанием миоглобина), напротив, осуществляют быстрые сильные сокращения.
3. Теплообразование и эффективность. Выделение мышцей тепла пропорционально силе и продолжительности изометрического тетануса; дополнительное тепло выделяется, если мышца укорачивается, совершая при этом работу. Эффективность в расчете на целую мышцу составляет 20-30%, эффективность ее сократительного аппарата — 40-50%.
Видео физиология мышц и мышечного сокращения - профессор, д.м.н. П.Е. Умрюхин
- Рекомендуем ознакомиться далее "Строение, функции и сокращение гладкой мускулатуры - с точки зрения физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 17.10.2024