
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Нейрорегуляция мышечной силы - с точки зрения физиологии человека
Оглавление статьи:- Частота потенциалов действия и тетаническое сокращение
- Регуляция сократительной силы двигательных единиц
- Электромиографическая диагностика
- Мышечная гипертрофия и атрофия
- Коротко
- Список использованной литературы
а) Частота потенциалов действия и тетаническое сокращение. При высокой частоте потенциалов действия возникает слитное устойчивое сокращение — тетанус; во время тетанического сокращения цитоплазматическая концентрация Са2+ повышается.
1. Произвольные сокращения. Силу своих скелетных мышц человек может изменять произвольно. Регуляцию силы осуществляет центральная нервная система. Прежде чем перейти к механизмам регуляции, рассмотрим, как зависит от частоты стимулов саркоплазматическая концентрация Са2+ и сокращение скелетной мышцы.
2. Кальциевые сигналы при одиночном сокращении и тетанусе. В изолированном волокне скелетной мышцы можно зарегистрировать люминесценцию чувствительного к Са2+ вещества-индикатора в качестве показателя изменений концентрации Са2+ во время изометрических сокращений (рис. 1).
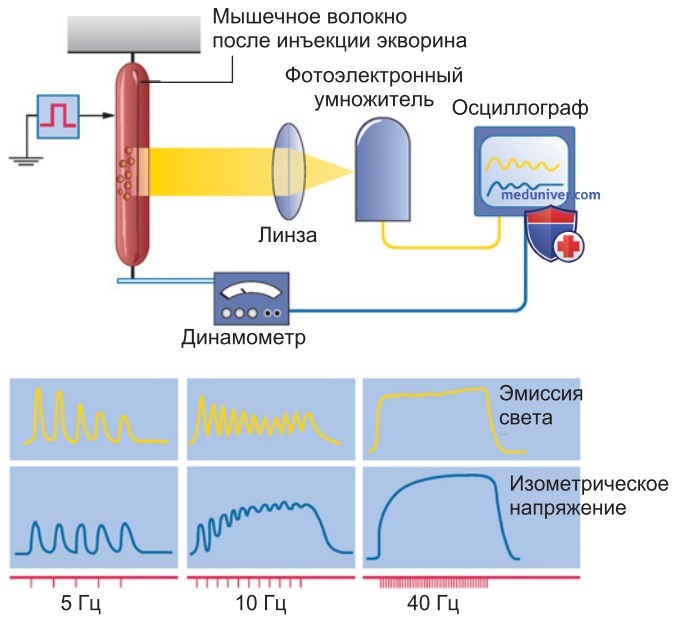
При стимуляции волокна с частотой 5 Гц высвобождаемый Са2+ тут же возвращается в СР и люминесцентные сигналы воспроизводят форму каждого отдельного мышечного сокращения. При частоте 10 Гц последовательные сокращения накладываются друг на друга, общее мышечное напряжение нарастает; происходит суперпозиция (суммация) одиночных сокращений.
Однако после каждого отдельного сокращения концентрация Са2+ в цитоплазме снова падает почти до уровня покоя. Только при еще более высокой частоте стимулов (и соответственно потенциалов действия), достигающей 20 Гц и больше, повышенная концентрация Са2+ удерживается в течение всего периода стимуляции, поскольку Са2+-АТФаза не успевает возвращать Са2+ в СР.
Теперь одиночные сокращения сливаются — сначала частично, затем полностью (рис. 1); возникает гладкий тетанус.
3. Частота стимуляции, обеспечивающая слитное сокращение — тетанус. Последовательные одиночные сокращения сливаются в гладкий тетанус, когда промежутки между стимулами (вызывающими потенциалы действия) составляют менее трети или четверти продолжительности одиночного сокращения. Кроме того, частота стимуляции, необходимая для развития тетануса, тем ниже, чем продолжительнее одиночное сокращение.
Следовательно, для тетанического сокращения волокон медленного типа достаточна более низкая частота, чем для волокон быстрого типа. При этом минимальный промежуток между последовательными стимулами в составе тетануса должен быть не короче рефрактерного периода, длительность которого примерно соответствует потенциалу действия (2-3 мс).
P.S. Тетанус-контрактура-тетания. Продолжительное сокращение в отсутствие потенциалов действия (например, при добавлении кофеина в среду) называется контрактурой. Это состояние следует отличать от тетануса, так же как и тетанию — следствие нарушения возбудимости мембран при дефиците Са2+ в среде. Особое внимание стоит уделять столбняку (tetanus); это заболевание, вызываемое токсином бактерии Clostridium tetani, характеризуется судорогами, опасными для жизни.
б) Регуляция сократительной силы двигательных единиц. Сила мышечных сокращений регулируется центральной нервной системой путем изменения частоты разрядов, генерируемых мотонейронами, и количества вовлекаемых в возбуждение двигательных единиц.
1. Сила сокращений и частота потенциалов действия. Как показано на рис. 1, сила тетанического сокращения зависит от частоты электрических сигналов. Это обстоятельство имеет физиологическое значение для нашего организма, поскольку произвольные сокращения обычно носят тетанический характер. При повышении частоты потенциалов действия мотонейрона с 10 до 50 в секунду (для мотонейронов некоторых мышц быстрого типа частота может составлять 100 Гц) возникает гладкий устойчивый тетанус (вместо неполного зубчатого тетануса) и сила сокращения повышается в 2-8 раз.
Даже при низкой частоте разряда суммарное напряжение мышцы удерживается на одном и том же уровне благодаря тому, что разные двигательные единицы достигают максимального сокращения асинхронно. Причины повышенного мышечного напряжения во время гладкого тетануса могут быть следующими:
- длительность сокращения, при которой степень растяжения последовательных эластичных элементов достаточна для передачи максимальной силы к сухожилиям;
- насыщение тропонина С ионами Са2+, для чего необходима высокая частота возбуждения.
2. Вовлечение двигательных единиц. Сила, развиваемая индивидуальной двигательной единицей при одиночном сокращении, практически не варьируется: все ее волокна либо сокращаются, либо расслаблены (поскольку каждое из них подчиняется закону все или ничего). Однако сила сокращений (а также скорость) скелетных мышц весьма эффективно изменяется за счет активации разного числа двигательных единиц. При слабом произвольном сокращении мышцы потенциалы действия наблюдаются лишь в некоторых двигательных единицах (рис. 2).
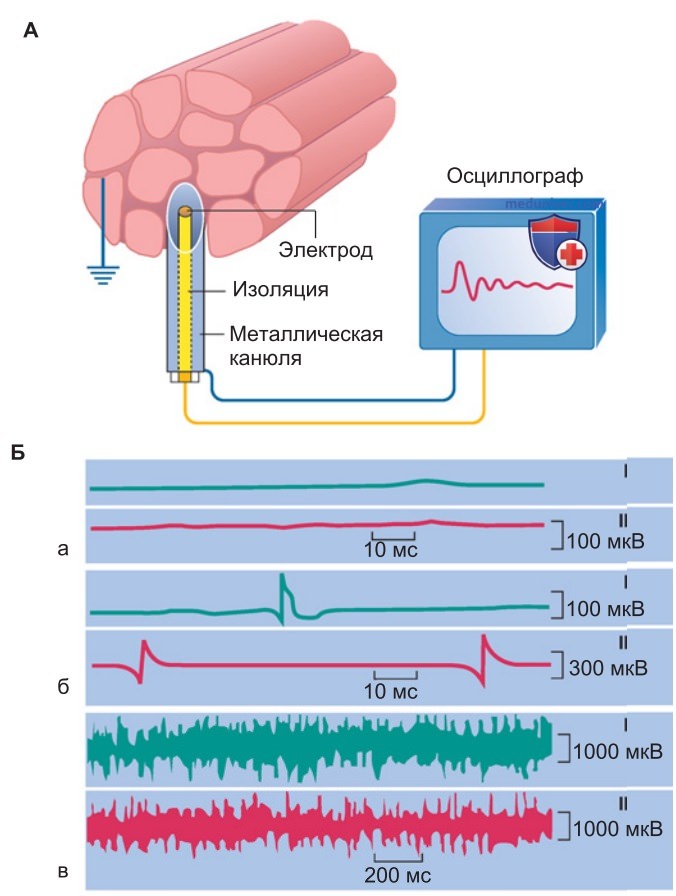
При сильном произвольном сокращении разряды возникают во многих двигательных единицах. В результате усиливается и регистрируемая с поверхности кожи интегральная электрическая активность мышцы — тем значительнее, чем интенсивнее сокращаются участки мышц под электродами, наложенными на кожу. Чем меньше размеры (а значит, и сила) каждой двигательной единицы, тем тоньше регулируется суммарное усилие всей мышцы.
3. Рефлекторный тонус. Даже в состоянии видимого покоя в некоторых мышцах регистрируется слабая электрическая активность. Низкочастотные разряды небольшого числа двигательных единиц могут обеспечивать непроизвольное рефлекторное напряжение позных мышц. Такой нейрогенный тонус модулируется системой γ-эфферентов мышечных веретен. Во время умственного напряжения или эмоционального возбуждения нейрогенный тонус непроизвольно усиливается, а полностью исчезает только при глубоком расслаблении.
в) Электромиографическая диагностика. Регистрация электромиограммы (ЭМГ) осуществляется в диагностических целях при подозрении на нервно-мышечное заболевание.
1. Клиническая электромиография. Регистрация ЭМГ позволяет прослеживать потенциалы действия двигательных единиц во время мышечной деятельности (рис. 2). С помощью поверхностных накожных электродов можно регистрировать суммарную активность мышц; игольчатые электроды, введенные в мышцу, обеспечивают более точные диагностические характеристики электрических сигналов. В обоих случаях частота и амплитуда потенциалов регистрируются внекле-точно. Амплитуда зависит от количества активных (генерирующих импульсные разряды) двигательных единиц или мышечных волокон в непосредственной близости от электродов.
Если активны несколько ближайших двигательных единиц, то их асинхронная активность создает картину повышенной частоты потенциалов действия. Электромиограмма дает также представление о том, сколько функциональных двигательных единиц содержат мышцы, активность которых регистрируют электроды.
2. Электромиографическое исследование при миотонии. Возбудимость сарколеммы повышается настолько, что даже при введении в мышцу игольчатого электрода спонтанно запускаются интенсивные разряды. Когда после периода покоя пациент произвольно напрягает мышцу, возникают продолжительные следовые разряды. Измененные ЭМГ-сигналы можно регистрировать также при нарушениях иннервации мышц. На первой стадии денервации (которая предшествует инактивационной атрофии мышцы) появляются спонтанные потенциалы действия (потенциалы фибрилляции).
После продолжительной, полностью завершившейся денервации, например при полиомиелите, атрофированные мышечные волокна замещаются соединительной тканью; у оставшихся сохранившихся электромиографических сигналов очень низкая амплитуда.
г) Мышечная гипертрофия и атрофия. Гипертрофия или атрофия могут сопровождаться долгосрочными изменениями мышечной силы.
1. Мышечная гипертрофия. Чем толще мышца, т.е. чем больше суммарная площадь поперечного сечения ее волокон, тем более значительную силу она может развить. Как известно, с помощью тренировки человек может добиться мышечной гипертрофии; при этом увеличивается диаметр мышечных волокон, тогда как их количество в мышце не изменяется (в случае гиперплазии количество мышечных волокон, напротив, возрастает!). В клетках гипертрофированной мышцы разрушается большее количество белка, чем синтезируется.
2. Мышечная атрофия. Если, наоборот, в течение длительного времени разрушение мышечных белков преобладает над их синтезом, наступает мышечная атрофия; развиваемая сила будет ниже нормы. Усиливающаяся атрофия характерна для бездействующих мышц, денервации, а также наблюдается при старении.
д) Коротко. Регуляция мышечной силы. Повышение частоты стимулов приводит к суммации одиночных сокращений, так что развивается неполный (зубчатый) тетанус (физиологическая форма сокращения!), вплоть до полного (гладкого) тетануса. При тетанических сокращениях цитоплазматическая концентрация Са2+ во время промежутков между импульсами остается повышенной.
При переходе от низкочастотного неполного тетануса к гладкому мышечная сила возрастает в 2-8 раз. ЦНС может регулировать мышечную силу посредством двух разных механизмов: изменением частоты разряда мотонейронов или вовлечением двигательных единиц.
При диагностике нарушений нервно-мышечных функций используется электромиографический метод.
Вследствие гипертрофии или атрофии мышц могут возникать стойкие изменения мышечной силы.
Видео физиология мышц и мышечного сокращения - профессор, д.м.н. П.Е. Умрюхин
- Рекомендуем ознакомиться далее "Механика сокращения скелетной мышцы - с точки зрения физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 17.10.2024