
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Слои эпидермиса и его строение
Одна из основных видимых частей кожи — многослойный ороговевающий эпидермис. Эпидермис — постоянно обновляющаяся структура, что позволяет расти его производным, называемым придатками (волосы и сальные железы, ногти и потовые железы). Толщина эпидермиса составляет от 0,4 до 1,5 мм, толщина всех слоев кожи — от 1,5 до 4,0 мм. Основная часть клеток эпидермиса — кератиноциты, объединены в четыре слоя, названных либо по расположению, либо по структурным свойствам клеток.
Эти клетки постепенно дифференцируются от пролиферативных базальных клеток, прилегающих к базальной мембране эпителия, до окончательно дифференцированных, ороговевших клеток, формирующих внешний слой и барьер кожи.
В различных слоях среди кератоцитов можно встретить отростчатые клетки двух типов — клетки Лангерганса и меланоциты. Кроме того, в базальном слое имеются особые чувствительные клетки Меркеля. В норме другие клетки, например лимфоциты, встречаются в эпителии кратковременно и достаточно редко. Существует множество регионарных различий эпидермиса и его производных. Некоторые из этих различий очевидны, такие, как толщина (например, кожа на конечностях и кожа на туловище), другие видны только на микроскопическом уровне.
Патологические изменения эпителия могут возникать в результате воздействия целого ряда факторов: повторяющихся механических травм (например, лихеноидный зудящий дерматит), воспаления (атопический дерматит и плоский лишай), инфекции (например, простая бородавка), нарушений в работе иммунной системы и цитокинов (как при псориазе), образования аутоантител (как при пузырчатке и буллезном пемфигоиде) или генетических дефектов, влияющих на дифференцировку, или структурные белки [как при врожденном буллезном эпидермолисе (ВБЭ), эпидермолитическом гиперкератозе, ихтиозе и при болезни Дарье].

Эпидермис—многослойный ороговевающий эпителий. Глубокий слой состоит из базальных клеток, которые лежат на базальной мембране (БМ), отделяющей соединительную ткань от базального слоя (БС).
Далее эти клетки дифференцируются в клетки шиповатого слоя (ШС), характеризующиеся большим количеством выростов и десмосом.
Шиповатые клетки постепенно превращаются в клетки зернистого слоя (ЗС), вырабатывающие многие компоненты рогового слоя.
В итоге постепенная дифференцировка кератиноцитов приводит к потере ими ядер и превращению в чешуйки, составляющие роговой слой (PC) — переплетение белков и гликолипидов.

Слои эпидермиса
а) Базальная мембрана эпидермиса. Кератиноциты — клетки эктодермального происхождения, которые являются основным типом клеток в эпителии. На их долю приходится не менее 80% общего количества клеточных элементов. Конечная судьба этих клеток — стать компонентом эпидермального барьера в качестве рогового слоя. Таким образом, значительную часть функций эпителия можно понять, изучив строение и развитие кератиноцитов.
Дифференцировка кератиноцитов (кератинизация) — генетически запрограммированная, тщательно регулируемая последовательность сложных морфологических и метаболических изменений, конечной точкой которых является полностью дифференцированные мертвые кератиноциты (корнеоциты), которые содержат кератиновые филаменты, матриксный белок и богатую белком плазматическую мембрану, поверхность которой связана с липопротеидами.
Кератины относятся к семейству фибриллярных белков и специфичны для всех эпителиальных клеток, в том числе кератиноцитов. Они играют структурную роль в клетках. У человека выявлено 54 гена, кодирующих кератины (34 эпителиальных и 17 волосяных кератинов). Совместная экспрессия определенной пары кератина зависит от типа клеток, типа ткани, стадии развития, степени дифференцировки и заболевания. Кроме того, важная роль данных молекул подчеркивается многочисленными проявлениями заболеваний, возникающих из-за мутаций в этих генах.
Таким образом, знания о синтезе кератина, его регуляции и структуре позволяют получить представление о дифференцировке эпидермиса и его строении.
Базальный слой (ростковый) содержит митотически активные кератиноциты цилиндрической формы, которые прикреплены нитями кератина (К5 и К14) к базальной мембране при помощи полудесмосом, а с окружающими клетками соединены десмосомами, что обеспечивает рост клеток в более поверхностных, дифференцированных слоях эпидермиса. Связанные с мембраной вакуоли, которые содержат пигментные меланосомы, переносятся из меланоцитов фагоцитами. Пигмент, находящийся в меланосомах, вносит свой вклад в общую пигментацию кожи, которая видна невооруженным глазом. Базальный слой — основное местонахождение митотически активных клеток эпидермиса. Результаты клеточно-кинетических исследований подтверждают, что базальный слой содержит клетки различной пролиферативной активности (стволовые клетки, временно амплифицированные клетки и дифференцированные клетки), а в исследованиях in vivo и in vitro было показано существование долгоживущих эпидермальных стволовых клеток.
Так как клетки базального слоя можно культивировать и использовать для восстановления эпидермиса и покрывать ими кожу обожжённых пациентов, такая популяция клеток считается содержащей долгоживущие стволовые клетки, которые обладают значительным пролиферативным потенциалом. Подобные клетки расположены в базальном слое и утолщениях волосяных фолликулов. Второй тип клеток, дочерних по отношению к стволовым, образуется в результате их нечастого деления, как симметричного, так и ассиметричного. Эти клетки продвигаются к поверхности кожи и дифференцируются. Эти клетки обеспечивают основное число делений, необходимое для стабильного самообновления, и являются наиболее распространенными клетками базального слоя. После прохождения нескольких циклов деления эти клетки дают начало третьему классу базальных клеток эпидермиса, постмитотическим клеткам, которые подвергаются окончательной дифференцировке. У человека в норме, от потери базальной клеткой контакта с базальным слоем до ее попадания в роговой слой, проходит не менее 14 дней. Переход в роговой слой и последующая десквамация занимают еще 14 дней.
При гиперпролиферативных или замедляющих рост состояниях эти периоды могут изменяться.
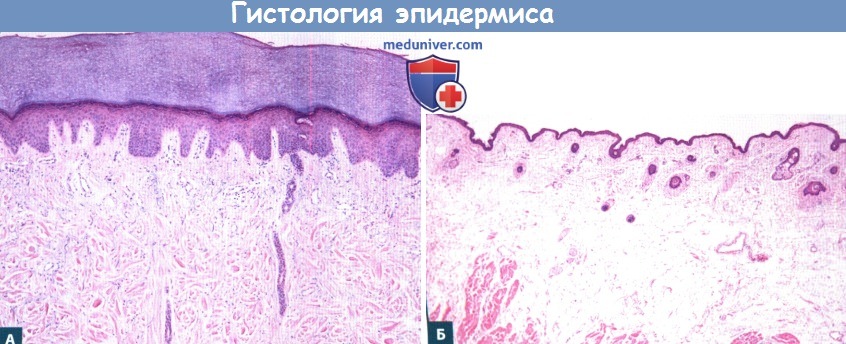
А. Кожа конечностей.
Б. Кожа век. Следует отметить, что эпидермис кожи конечностей значительно толще за счет компактного слоя,
а также более глубоких слоев эпидермиса.
б) Шиповатый слой. Форма, структура и содержимое шиповатых клеток зависят от их расположения в среднем слое эпидермиса. Свое название клетки получили в связи с шиповидной формой краев клеток на гистологическом срезе. Супрабазальные шиповидные клетки имеют многогранную форму с круглыми ядрами. В результате дифференцировки и продвижения вверх через эпидермис, клетки становятся все более плоскими и в них выявляются органеллы, известные как «пластинчатые гранулы» (см. «Зернистый слой»). Шиповатые клетки также содержат большие скопления кератиновых филаментов, расположенных вокруг ядра и входящих в состав периферических десмосом клетки.
Шиповатые клетки задерживают устойчивые К5/К14 кератины, вырабатываемые в базальных слоях, без гиперпролиферативных нарушений не синтезируют новые молекулы информационной РНК (иРНК) для данного белка. В этом слое происходит синтез новых кератиновых пар К1/К10. Эти кератины характерны для эпидермального типа дифференцировки и поэтому называются «специфическими для дифференцировки или специфическими для кератинизации кератинами». При таких заболеваниях, как псориаз, актинический кератоз и при заживлении ран, синтез К1 и К10 мРНК уменьшается, тогда как синтез и трансляция иРНК Кб и К16 возрастают. С такими отклонениям в экспрессии кератина связано нарушение нормальной дифференциации в зернистом и ороговевшем эпидермальном слоях (см. «Зернистый слой» и «Роговой слой»).
Информационная РНК для К6 и К16 обычно присутствует на всем протяжении эпидермиса, однако информация транслируется только при стимуляции пролиферации.
«Шипы» шиповатых клеток богаты десмосомами, модификация кальций-зависимой клеточной поверхности которых способствует адгезии эпидермальных клеток и оказанию сопротивления механическому воздействию. Несмотря на то что десмосомы относятся к адгезивным соединениям, последние ассоциированы с актиновыми микрофиламентами поверхностями межклеточных контактов через определенный набор кадгеринов (например, Е-кадгерины) и внутриклеточные молекулы адаптера катенина. Десмосомы являются интегральными медиаторами внутриклеточной адгезии, что было отчетливо продемонстрировано при заболеваниях с нарушениями этих структур.
Важное значение кальция как медиатора адгезии видно на примере заболеваний, проявляющихся характерной несостоятельностью эпидермальных контактов: болезни Дарье (фолликулярный дискератоз) и болезни Хейли — Хейли (доброкачественный (хронический) семейный пемфигус). Оба заболевания вызваны мутациями в генах, регулирующих транспорт кальция, SERCA2 при болезни Хейли-Хейли.
В этом слое эпидермальных клеток также образуются ламеллярные гранулы. Эти секреторные органеллы доставляют предшественники липидов рогового слоя в межклеточное пространство. Генетические заболевания служат доказательством существенного значения стероидного и липидного метаболизма в процессе отторжения ороговевших клеток. Например, мутация стероидной сульфатазы приводит к ретенции гиперкератоза при рецессивном Х-сцепленном ихтиозе.
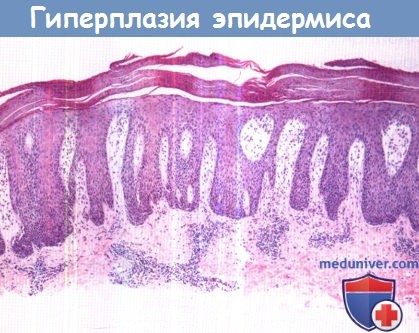
Гиперпролиферация эпидермиса проявляется такими заболеваниями, как псориаз (фото),
лихеноидный зудящий дерматит, атопический дерматит, красный плоский лишай, бородавки и сыпь.
в) Зернистый слой эпидермиса. Названный так из-за базофильных кератогиалиновых гранул, выделяющихся в клетке в этом слое эпидермиса, зернистый слой является местом зарождения структурных клеток, формирующих эпидермальный барьер, а также некоторых белков, участвующих в этом формировании. Первоначально кератогиалиновые гранулы состоят из профилагрина, кератиновых филаментов и лори-крина. Именно в этом слое начинается формирование оболочки роговой клетки. В конечном счете филагрин вырождается в молекулы, включающие уроканиновую и пироглутаминовую кислоты, которые способствуют гидратации рогового слоя и участвуют в фильтрации ультрафиолетовых лучей. Лорикрин — белок, обогащенный цистеином. Этот белок представляет основную часть белкового компонента роговой оболочки. После освобождения от кератогиалиновых гранул лорикрин связывается с десмосомальными структурами элафи-ном и энвоплакином и впоследствии прикрепляется к цитоплазматической мембране с помощью клеточных трансаминаз (TGMs, первичная TGMs 3 и 1), формируя, таким образом, оболочку роговых клеток.
Было показано, что мутации в гене TGM1 являются причиной некоторых форм ламеллярного ихтиоза. Другая форма ихтиоза — вульгарный ихтиоз- вызвана мутациями в гене филагрина. Аномалии лорикрина приводят к синдрому Фовинкеля с ихтиозом и ложным спонтанным дактилолизом, а также к симметричной прогрессирующей кератодермии. Эти данные подчеркивают значение правильного формирования роговой оболочки в нормальной кератинизации эпидермы.
Заключительный этап дифференцировки клеток зернистого слоя состоит в запрограммированной гибели клеток.
В ходе этого процесса зернистые клетки становятся окончательно дифференцированными корнеоцитами, их апоптоз вызывает разрушение ядра и всех клеточных включений, кроме кератиновых филаментов и матрикса филагрина.
г) Роговой слой эпидермиса. Полная дифференцировка клеток зернистого слоя приводит к формированию наложенных друг на друга рядов безъядерных, уплощенных, ороговевших клеток, образующих роговой слой. Последний осуществляет механическую защиту кожи и является барьером, с одной стороны, предотвращающим потерю организмом воды, а с другой, ограничивающим проникновение растворимых веществ из окружающей среды. Барьер рогового слоя образован двумя компонентами: лишенными липидов и богатыми белками (1) корнеоцитами, окруженными (2) экстрацеллюлярным липидным матриксом. Эти два компонента обеспечивают ряд отчасти изолированных, однако дополняющих друг друга функций, совокупность которых определяет «барьерную активность» эпидермиса. Первичными функциями экстрацеллюлярного липидного матрикса являются регуляция проницаемости, десквамация, противомикробный эффект, выведение токсических веществ. В свою очередь корнеоциты обеспечивают механическую прочность рогового слоя, увлажнение, инициацию опосредованного цитокинами воспаления, защиту от УФ излучения.
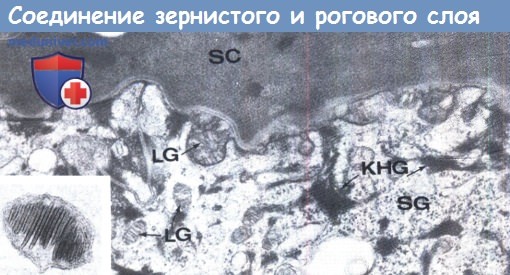
Ламеллярные гранулы (lamellar granules — LG) в межклеточном пространстве и цитоплазме рогового слоя.
Отчетливо видны кератогиалиновые гранулы (KHG). Вставка: Ламеллярная гранула х28,750.

- Рекомендуем далее ознакомиться со статьей "Клетки эпидермиса некератиноцитовые"
Оглавление темы "Анатомия кожи.":- Подкожная клетчатка и ее патология
- Функции кожи и обзор ее строения
- Слои эпидермиса и его строение
- Клетки эпидермиса некератиноцитовые
- Дермально-эпидермальное соединение кожи
- Слои дермы и их строение
- Кровеносные сосуды кожи и их строение
- Лимфатические сосуды кожи и их строение
- Нервы и рецепторы кожи
- Гиподерма и ее строение