
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Механизм миграции эозинофил в ткани
Избирательное рекрутирование эозинофилов к очагам воспаления является, в основном, функцией цитокинов, активирующих эозинофилы; цитокинов, активирующих эндотелий, и хемокинов. Представители суперсемейства генов селектинов, интегринов и иммуноглобулинов аналогично другим лейкоцитам обеспечивают сигнальные пути, задействованные в миграции эозинофилов.
В особенности это касается постоянно экспрессируемого эозинофилами интегрина VLA-4, взаимодействующего с лигандом VCAM-1, индукцию которого на эндотелиальных клетках обеспечивают, прежде всего, Th2-цитокины (ИЛ-4 и ИЛ-13). После проникновения сквозь сосудистую стенку эозинофилы связываются с белками экстрацеллюлярного матрикса. Здесь такие поверхностные факторы, как b2-интегрины, например, CD11 b/CD18 (Мас-1), связываются с фибриллярными белками, например, фибронектином, ламинином, коллагеном, и при этом не только определяют, где эозинофилы будут локализоваться, но и, по всей видимости, продлевают срок их жизнедеятельности. В этом плане принципиальное значение для эффекторной функции эозинофилов (включая дегрануляцию) также имеет интегрин CD11b/CD18 (Мас-1).
а) Цитокины активирующие эозинофилы. Цитокины, активирующие эозинофилы, продуцируются многими видами клеток (за исключением Т-клеток и тучных клеток). К их числу относят кератиноциты, эндотелиальные клетки, моноциты, а также сами эозинофилы. Цитокины, активирующие клетки (ИЛ-3, ИЛ-5, ГМ-КСФ и др.), усиливают хемотаксический ответ, а также определяют и другие многочисленные эффекты, включая созревание и выживание клеток, продукцию лейкотриенов.
б) Цитокины активирующие эндотелий. Во время миграции эозинофилов происходит по меньшей мере три разных типа эндотелиальной активации. Первый тип — это экспрессия Р-селектина, которая происходит, когда тельца Вейбеля-Паладе, содержащиеся в эндотелиальной клетке, чрезвычайно быстро транспортируются на клеточную поверхность после воздействия на эндотелиальную клетку гистамина, лейкотриенов и ряда других воспалительных медиаторов.
Экспрессия Р-селектина на поверхности эндотелия инициирует роллинг лейкоцитов (через PSGL-1), который представляет собой важный этап перед плотной адгезией и миграцией через эндотелий. Второй тип активации эндотелия индуцируется неспецифическими активаторами, такими как ИЛ-1 и ФНО-а. Эти цитокины стимулируют экспрессию на эндотелии Е-селектина, ICAM-1 и VCAM-1, а также индуцируют продукцию хемокинов эндотелиальными клетками. Третий тип активации эндотелия инициируется интерлейкинами ИЛ-4 и ИЛ-13.
Эти цитокины избирательно индуцируют VCAM-1, которая играет центральную роль в привлечении к очагам аллергического воспаления VLА4-положительных клеток, включая эозинофилы, базофилы и лимфоциты.
в) Хемокины. Переход от роллинга к стабильно фиксированной форме существенным образом облегчается под действием лигандов CCR3 — СС хемокинов. Индукция экспрессии хемокинов активированными эндотелиальными клетками приводит к более высоким концентрациям хемокинов на поверхности эндотелия или около нее, которые временно воздействуют на авидность интегринов β1 и β2; что приводит к плотной адгезии эозинофилов на клетке эндотелия.
Экспрессия хемокинов тканями создает сигнальный градиент, который ведет эозинофилы в толщу ткани. CCL11 проводит эозинофилы туда, где они и должны быть в норме — в тимус, матку, молочные железы и желудочно-кишечный тракт. При заболеваниях ассоциированных с патологией Th2 звена, цитокины этой группы лимфоцитов индуцируют экспрессию хемокинов. В легких больного бронхиальной астмой и эозинофильной пневмонией увеличено число CCL11, CCL24 и CCL26, обнаруживаемых там наряду с другими хемокинами, такими как лейкотриен В4 и галектин-9.
В желудочно-кишечном тракте CCL26 играет ключевую роль в развитии эозинофильного эзофагита, в то время как CCL11 вовлечен в эозинофильное поражение нижних отделов пищеварительного тракта. В коже ИЛ-4, Ил-13 и ФНО-а стимулируют продукцию CCL11, CCL24, CCL26 тучными клетками и лимфоцитами, а также CCL11, CCL26 фибробластами и CCL26 кератиноцитами. Как и при эозинофильном эзофагите, CCL26 может играть роль в патогенезе атопического дерматита, при котором сывороточный уровень данного хемокина коррелирует с активностью заболевания.
Метаболиты арахидоновой кислоты, в частности цистеиниловые лейкотриены, лейкотриены С4, D4, Е4 и простогландин-2 задействованы в передвижении эозинофилов, что подтверждается способностью антагонистов рецепторов лейкотриенов уменьшать эозинофилию в легких и крови, а также тем, что у мышей с деплецией рецепторов лейкотриена В4 наблюдается достоверное уменьшение концентрации эозинофилов в легких после воздействия аллергена. Перемещение в ткани эозинофилов, базофилов и Th-клеток происходит посредством рецепторов CRTH2 (CD294) — высокоаффинных рецепторов простогландина-2 второго типа.
Идентифицируют и другие факторы, оказывающие влияние на перемещение эозинофилов. Так эозинофилы экспрессируют в высоких концентрациях рецепторы гистамина 4 типа, через которые осуществляется хемоаттракция и активация. Белок внеклеточного матрикса, периостин, закодированный в гене, который подвергается индукции ИЛ-13, облегчает инфильтрацию эозинофилов в ткани за счет прямого влияния на адгезию; периостин синтезируется в больших количествах при эозинофильном эзофагите, а его концентрация коррелирует с числом эозинофилов в биопсии.
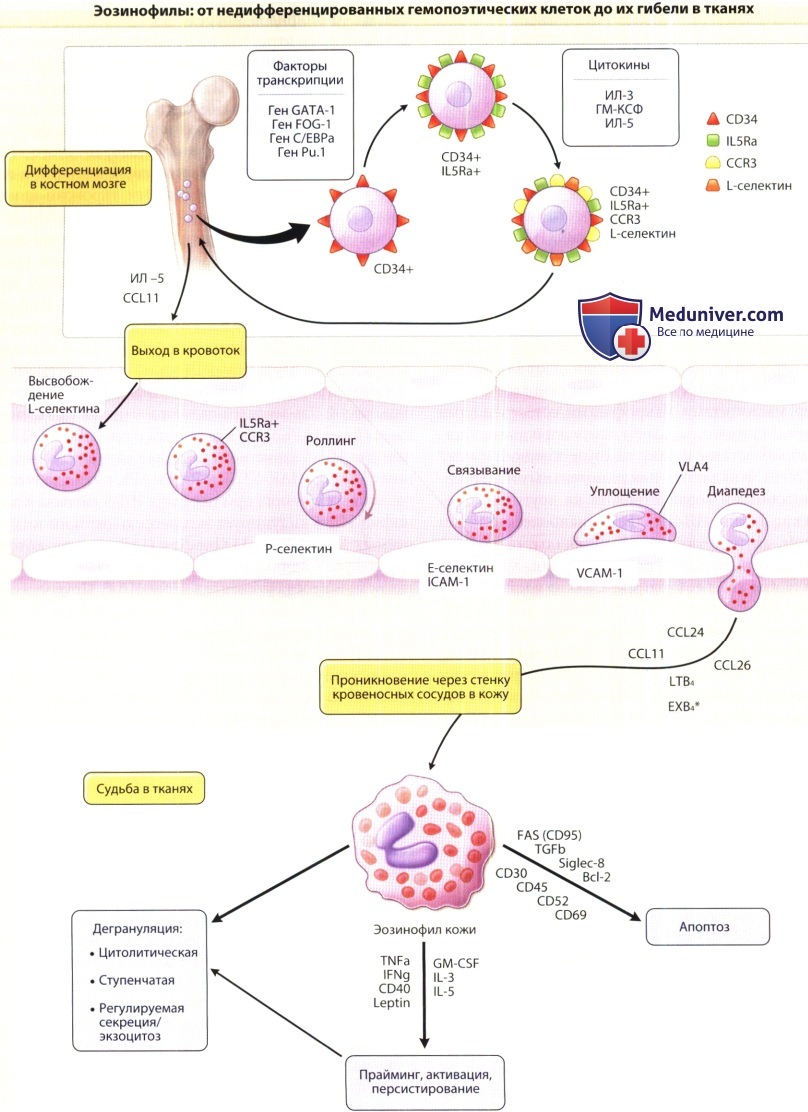
На рисунке представлен жизненный цикл эозинофилов от дифференцировки в костном мозге к трансвазальной миграции и гибели в тканях (приведены ключевые факторы, участвующие в этом процессе).
- Рекомендуем далее ознакомиться со статьей "Механизм ативации эозинофил"
Оглавление темы "Эозинофилы при болезнях кожи.":- Происхождение и развитие эозинофилов
- Цитокины и сигнальные пути эозинофилов
- Cтруктура эозинофил и состав эозинофильных гранул
- Функции эозинофил и их роль в иммунитете
- Компоненты эозинофил и их функции
- Состав и компоненты кристаллов Шарко-Лейдена эозинофил
- Механизм миграции эозинофил в ткани
- Механизм ативации эозинофил