
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Состав и компоненты кристаллов Шарко-Лейдена эозинофил
а) Активные формы крислорода кристаллов Шарко-Лейдена. Активные формы кислорода являются важными медиаторами повреждения тканей эозинофилами. Активный кислород, синтезированный НАДФ оксидазой во время окислительного взрыва, индуцированного эозинофилами, представлен такими формами как супероксиды, гидроксильные радикалы и перекись водорода. Перекись водорода способна образовывать гипогалоидные кислоты за счет действия эозинофильной пероксидазы, которая сама по себе обладает цитотоксическим действием. Активные формы кислорода также способны усиливать воспалительный ответ, индуцируя экспрессию генов и пролиферацию Т-клеток.
б) Липидные медиаторы. Общая информация. Липидные тельца эозинофилов выполняют функцию хранилища арахидоновой кислоты. Эозинофилы способны выделять многочисленные метаболиты этой жирной кислоты, включая разные виды цистеинил-LT за счет 5-липоксигеназного пути (LTC4, LTD4, LTE4), а также тромбаксаны и проста-гландины за счет циклогеназного пути (тромбоксан В2, простагландин Е2 и простагландин F1).
в) Цитокины. Эозинофилы являются существенным источником факторов роста и регуляторных и про-воспалительных цитокинов и хемокинов. Различные виды факторов роста, синтезируемых эозинофилами включают трансформирующий фактор роста-а, трансформирующий фактор роста-у, фибробластный фактор роста-2, фактор роста сосудистого эндотелия, фактор роста нервов и тромбоцитарный фактор роста. Существуют данные, что все эти факторы роста способствуют стромальному фиброзу и утолщению базальной мембраны в участках хронического эозинофильного воспаления, к числу которых относятся полипы носа, бронхиальная астма и, скорее всего, атопический дерматит. Продукция и выделение фактора роста нервов эозинофилами вызывает воспаление нервной ткани. Еще одна группа цитокинов, выделяемых эозинофилами, модулирует другие иммунные клетки.
К таким цитокинам относятся фактор некроза опухоли-а, макрофагальный воспалительный белок (MIP) - 1а (CCL3), ИЛ-1а, ИЛ-2, ИЛ-3, ИЛ-4, ИЛ-5, ИЛ-6, CXCL8 (ИЛ-8), ИЛ-10, ИЛ-12, ИЛ-13, ИЛ-16, ГМ-КСФ, интерферон-у. Кроме того, в дополнение к CCL3 и CXCL8 эозинофилы синтезируют CXCL13 (хемоаттрактант В лимфоцитов), CCL5 (регулятор активации нормальный Т-клеток) и CCL11. Цитокин CCL5 накапливается очевидно в двух внутриклеточных компартментах:
(1) матриксе специфических эозинофильных гранул и (2) малых секреторных везикулах. Все эти цитокины в норме вырабатываются покоящимися эозинофилами в небольшом количестве, Однако в случае активации их рецепторов иммуноглобулинами, компонентами комплемента или циткоинами (в том числе и производимыми самими эозинофилами по аутокринному механизму в условиях воспаления — см. ниже) синтез всех этих веществ индуцируется. Важно учитывать, что эозинофилы производят три главных цитокина, играющих роль в их созревании и дифференцировке — (1) ИЛ-3,
(2) ИЛ-5 и (3) гранулоцитарно-макрофагальный колниестимулирующий фактор ГМ-КСФ. Кроме того, вырабатываются и хемокины задействованные в хемотаксисе CCL5 и CCL11. Известно, что синтез и секреция эозинофилами гранулоцитарно-макрофагального колониестимулирующего фактора осуществляется по пептидил-пропил изомераза-зависимому механизму. Хотя объем синтезированных эозинофилами хемокинов сравним с таковым у Т-клеток, относительный вклад эозинофильных цитокинов в процесс воспаления является весьма ограниченным. Обобщая сказанное, необходимо отметить, что цитокины эозинофилов могут выступать в качестве как аутокринных, так и паракринных агентов и играют патофизиологическую роль.
г) Экспрессия белков на поверхности эозинофилов. Эозинофилы синтезируют многочисленные рецепторы и другие факторы на поверхности своей мембраны и за счет этого взаимодействуют с внеклеточными окружением, при этом среди них нет ни одного уникального для эозинофилов рецептора. Они были идентифицированы как при помощи проточной цитометрии, так и функциональными методами, их можно сгруппировать следующим образом: хемотаксические факторы и рецепторы комплемента, включая хемокины, факторы активации тромбоцитов, лейкотриены; семейство иммуноглобулиновых супергенных мембранных рецепторов, в число которых входят иммуноглобулины; рецепторы цитокинов, включая описанные выше; рецепторы адгезии молекул; рецепторы, участвующие в апоптозе; а также вспомогательные рецепторы и факторы на поверхности клетки. Мембранные белки эозинофилов считаются многообещающей целью для терапевтического вмешательства.
д) Хемотаксические факторы и рецепторы комплемента. Хемотаксические факторы играют важную роль в управлении переходом клеток в очаг воспаления, равно как и в нахождении клетками своих участков в непатологических условиях (например, миграция эозинофилов в ткани желудочно-кишечного тракта). Эозинофилы имеют рецепторы к большому количеству хемотаксических агентов, таких как лейкотриен В4, фактор активации тромбоцитов, бактериальные антигены (N-формил-метиониллейцил-фелиал анин), анафилотоксин комплемента, С3 и С5а. Связывание хемокинов с соответствующими рецепторами на эозинофилах опосредованно влияет на многие биологические процессы. Помимо изменения формы клеток и миграции, происходят активация клеток, интернализация рецепторов, индукция респираторного взрыва и транзиторная активация адгезивности интегринов. Фактор активации тромбоцитов реализует свое действие через специфические рецепторы, которые были успешно клонированы.
Фактор активации тромбоцитов является одним из наиболее мощных хемоаттрактантов для эозинофилов и действует на них в значительно большей степени, нежели не нейтрофилы. Данный фактор также индуцирует выделение белков из гранул, активных форм кислорода и лейкотриена С4 из эозинофилов. Лейкотриен В4 через свои рецепторы стимулирует хемотаксис эозинофилов, усиливает метаболизм арахидоновой кислоты и увеличивает количество активных форм кислорода. Он не индуцирует дегрануляцию эозинофилов. Для эозинофилов также характерны репепторы к лейкотриену D4. Хемотаксины, перечисленные выше, оказывают мощное воздействие на эозинофилы, однако не являются специфичными и все влияют также и на другие виды лейкоцитов. Так как многие эозинофил-ассоциированные заболевания сопровождаются инфильтрацией тканей эозинофилами в отсутствие нейтрофилов, идентификация CCR3 рецепторов и их лигандов стала существенным прорывом в открытии эозинофил-селективных хемотаксинов.
Становится все более очевидным, что хемокиновый рецептор CCR3 и его основные лиганды (CCL5, CCL11, CCL13, CCL24, CCL26) играют решающую роль как в гомеостатическом, так и в связанном с воспалением привлечении эозинофилов в ткани. Неудивительно, что рецептор CCR3 обладает самым высоким уровнем экспрессии среди хемокиновых рецепторов у эозинофилов человека. CCR3 представляет собой семидоменный связанный G-субъединицей мембранный рецептор, способный передавать мощные сигналы, как позитивного, так и негативного характера внутрь клетки в зависимости от того, с каким лигандом взаимодействует.
е) Семейство мембранных иммуноглобулиновых супергенных рецепторов. Многие исследования функций эозинофилов, таких как фагоцитоз, антигензависмая цитотоксичность, окислительный метаболзм, продукция лейкотриенов С4 и выживаемости эозинофилов были выполнены при помощи IgG-покрытых мишеней. Среди рецепторов поверхности эозинофила наиболее экспрессируемыми оказались FcγRII (CD32), которые связывали агрегированные IgG, особенно подклассов IgG 1 и IgG3. Связывание этих иммуноглобулинов с рецепторами может играть потенциально значимую роль в дегрануляции эозинофилов при паразитарных и аллергических заболеваниях, наравне с другими эозинофильными функциями. Свежевыделенные эозинофилы из всех IgG рецепторов экспрессируют исключительно FcγRII (CD32), однако клетки можно стимулировать при помощи цитокинов, в частности интерферона-γ, после чего они начинают синтезировать FcγRI (CD64) и FcγRlII (CD16), a синтез FcγRII (CD32) усиливается.
FcαRI (CD89) рецептор IgA также представлен на мембране эозинофилов. Кроме того, эозинофилы обладают участками для связывания секторного компонента IgA, наиболее мощного стимулятора эозинофильной дегрануляции. Взаимодействие IgA с эозинофилами усиливается под дейсвтием Th2 цитокинов, ИЛ-4 и ИЛ-5. Эти наблюдения наряду с тем фактом, что многие эозинофилы обнаруживаются в эпителиальной ткани, например, желудочно-кишечного тракта, подтверждают значимую роль эозинофилов и секреторных IgA в формировании иммунной защиты слизистых.
Члены суперсемейства иммуноглобулинов являются трансмембранными молекулами типа I, которые имеют общие с иммуноглобулинами структурные особенности глобулярных доменов. Молекулы межклеточной адгезии (ICAM) -1 (CD54) и1САМ-3 (CD50) входят в это суперсемейство, экспрессируются на эозинофилах и, по всей видимости, играют важную роль в клеточной адгезии при взаимодействии лейкоцитов друг с другом, а также при контакте лейкоцитов с клетками тканей за счет функционального антигена лимфоцитов (LFA)-1 (aLβ2; CD 11а/CD18) в качестве контрлиганда.
ж) Рецепторы цитокинов. Рецепторы цитокинов присутствуют на поверхности эозинофилов в небольшом количестве. Легче всего обнаруживаются рецепторы ИЛ-3 (CD123), ИЛ-5 (CD125) и ГМ-КСФ (CD116), причем все они имеют общую β-цепь (CD132). Замечено, что эозинофилы активируются и рядом других цитокинов, несмотря на то, что рецепторы этих цитокинов трудно обнаружить методом проточной цитометрии. К таким рецепторам относятся рецепторы фактора стволовых клеток (рецептор c-kit; CD117), ИНФ-у (CD119), ФНО-a (CD120), ИЛ-4 (CD124), ИЛ-9 (CD129 и CD 132), ИЛ-13β (gp65), ИЛ-2 (CD25) и рецептор TGF-β.
з) Рецепторы адгезивных молекул. Рецепторы молекул межклеточной адгезии экспрессированы на клеточной поверхности эозинофилов и опосредованно участвуют в миграции эозинофилов в ткани, эти рецепторы также осуществляют общее взаимодействие между клетками.
Выделяют три группы таких рецепторов: (1) IgSF, селектины и их (2) гликопротеиновые контрлиганды, а также (3) интегрины. L-селектин (CD62L) и гликопротеиновый лиганд-1 Р-селектина (PSGL-1, CD162) имеют высокий уровень экспрессии, тогда как лиганды Е-селектина (например, сиалил-Льюис X (CD15s)] экспрессированы в очень низких концентрациях. Самой важной парой селектинов в процессе миграции эозинофилов в ткани являются Р-селектин и PSGL-1.
Эозинофилы экспрессируют на своей поверхности целый ряд интегринов (β1, β2 и β7), которые облегчают их адгезию к внеклеточным матричным протеинам, молекуле сосудистой клеточной адгезии 1 (VCAM1, CD106), расположенной на активированном эндотелии, или молекуле ICAM-1, которая находится на «дремлющем» или активированном эпителии, а также на активированном эндотелии. Интегрины представляют собой гетеродимеры, включающие а- и β-субъединицы, соединенные нековалентной связью. Интегрины β1, экспрессированные на эозинофилах, включают интегрин α4β1 (VLA-4), который связывается с молекулой VCAM-1, расположенной на активированном эндотелии, и внеклеточным матричным протеином, фибронектином. Адгезия эозинофилов с фибронектином индуцирует аутокринную продукцию цитокинов, увеличивающих выживаемость эозинофилов (ИЛ-3, ИЛ-5 и ГМ-КСФ). Еще один интегрин, экспрессированый на эозинофилах,— это интегрин α6β1 который опосредованно свя-зывывается с другим матричным протеином, ламинином.
На эозинофилах находятся четыре интегрина β2: aLβ2, (LFA-1), αMβ2, (Мас-1),аХ(3, и aDβ2. Эти интегрины связываются с ICAM-1,—2 и -3; VCAM-1; фибриногеном и фрагментом комплемента C3Bi. Наконец, эозинофилы также синтезируют α4β7, которые являются лигандом для адрессиновой молекулы клеточной адгезии слизистой кишечника первого типа (MAdCAM-1), которая, вероятно, играет роль в таксисе эозинофилов по отношению к слизистой желудочно-кишечного тракта. Кроме того, α4β7 способствует адгезии к фибронектину и васкулярной молекуле клеточной адгезии первого типа (VCAM-1).
и) Рецепторы, задействованные в процессе апоптоза. Эозинофилы экспрессируют несколько «рецепторов смерти», например, Fas-рецептор (CD95), Siglec-8, CD30, CD45, Campath (CD52) и CD69, а также важные внутриклеточные регуляторы апоптоза эозинофилов, такие как представители семейств В-клеточного лейкоза/лимфомы-2 (Вс1-2) и ингибиторов апоптоза. Заболевания, характеризующиеся эозинофилией, возникают как минимум отчасти из-за задержки или дефекта апоптоза, что вызывает накопление и персистирвоание эозинофилов в крови и/или тканях.
к) Факторы, действующие совместно. Различные продукты, синтезируемые эозинофилами в ответ на активацию рецепторов, не обязательно функционируют по отдельности, а скорее работают в тесной связи для реализации своих биологических эффектов. Например, совместное выделение фактора тканевого роста-αβ, фактора роста фибробластов-2, фактора роста сосудистого эндотелия, матриксной металлопротеиназы-9 и ее ингибитора усиливает пролиферацию фибробластов и продукцию белков межклеточного матрикса. Эозинофилы влияют на собственную продукцию и на продукты синтеза других клеток, например, обусловливают выделение тромбоцитами TGF-β. После интрадермального внедрения эозинофилов начинается продукция внеклеточных белков, включая тенасцин и проколлаген 1, а также упорядочивание миофибробластов. Эозинофил-индуцированный фиброз наблюдается в легких и сердце пациентов с гиперэозинофильным синдромом, внутри органа и вокруг него при других фиброзирующих/склерозирующих заболеваниях, а также в коже больных эозинофильным фасциитом (синдромом Шульмана), при синдроме эозинофильной миалгии и синдроме токсического масла.
Белки эозинофильных гранул, главный основный белок-1 и эозинофильный нейротоксин наряду с другими нейроактивными медиаторами, синтезируемыми эозинофилами (фактор роста нервов, вазоактивный интестинальный пептид и субстанция Р), оказывают влияние на функционирование нервной системы. Фактически эозинофилы и эозинофильные гранулы часто обнаруживаются рядом с нервными окончаниями. Эозинофил-индуцированная дисфункция нервных окончаний, вероятно, является важной частью нарушений моторики желудка, обнаруживаемых в исследованиях у лиц с пищевой аллергией, дисфункции парасимпатических мускариновых М2 рецепторов у астматиков, а также вносит вклад в формирование зуда и других патологических процессов у больных с атопическим дерматитом и прочими кожными заболеваниями. В целом, ответ эозинофилов на поверхностные факторы, определяет их роль при различных заболеваниях и в норме.
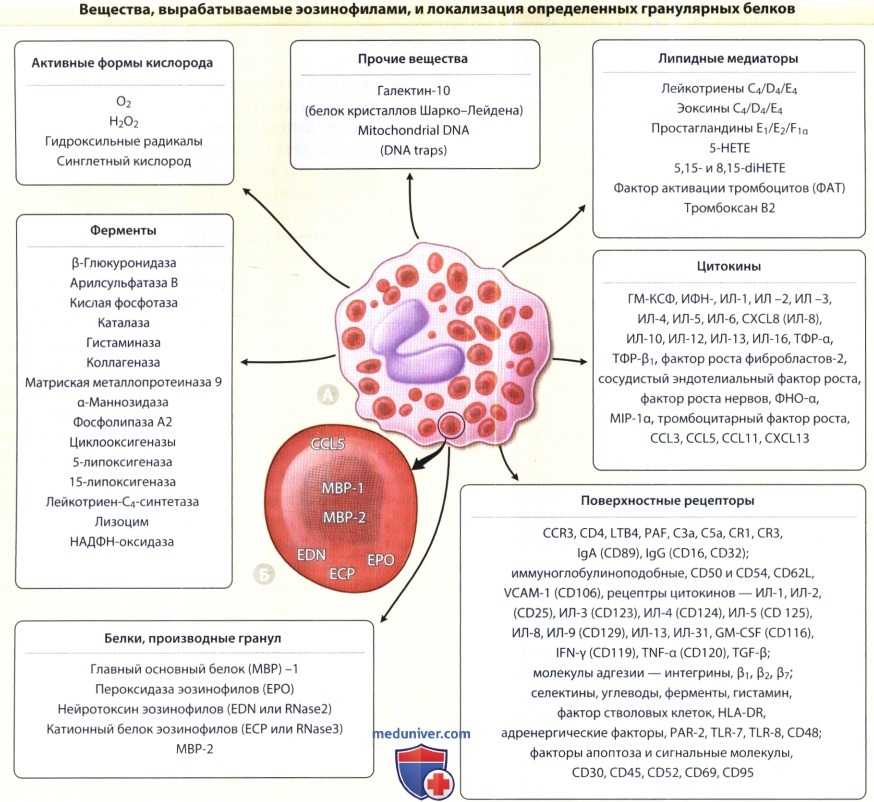
Эозинофилы вырабатывают множество веществ, среди которых цитотоксические белки гранул, играющие патогенетическую роль.
Характерные для эозинофилов гранулы крупнозернистые и, как следует из их названия, эозинофильные, т.е. окрашиваются эозином.
Определенные белки гранул располагаются в ядре и матриксе специфических цитоплазматических гранул.
А. Интактный эозинофил дермы с типичными гранулами и характерным двудольным ядром.
Б. Типичные специфические (вторичные) гранулы эозинофилов с электроноплотным кристаллическим ядром и прозрачным матриксом, что связано с характерной локализацией определенных гранулярных белков.
- Рекомендуем далее ознакомиться со статьей "Механизм миграции эозинофил в ткани"
Оглавление темы "Эозинофилы при болезнях кожи.":- Происхождение и развитие эозинофилов
- Цитокины и сигнальные пути эозинофилов
- Cтруктура эозинофил и состав эозинофильных гранул
- Функции эозинофил и их роль в иммунитете
- Компоненты эозинофил и их функции
- Состав и компоненты кристаллов Шарко-Лейдена эозинофил
- Механизм миграции эозинофил в ткани
- Механизм ативации эозинофил