
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Химическая синаптическая передача (возбуждение, торможение) - с точки зрения физиологии человека
Оглавление статьи:- Возбуждающие синапсы
- Двигательная концевая пластинка
- Тормозные синапсы
- Тормозные ионные токи
- Коротко
- Список использованной литературы
В 1812 г. британский первооткрыватель Чарльз Уотертон писал о своем путешествии по Гайане: «Местный охотник выстрелил в обезьяну, сидевшую на дереве прямо над ним. Он промахнулся, и стрела при падении задела его руку.
Предчувствуя свой конец, охотник лег на землю, попрощался с товарищами и умер». Яд «вурали» (кураре), которым аборигены смазывали стрелы, Уотертон привез в Англию и через несколько лет вместе с врачом Броди опубликовал сообщение: «После подкожного введения кураре у осла отнялись ноги. Его дыхание стали поддерживать через трахею с помощью воздуходувных мехов.
Через 2 ч осел поднялся, но когда прекратили искусственное дыхание, снова упал. Дальнейшая поддержка дыхания позволила восстановить нормальное состояние животного». Яд Уотертон продолжал хранить.
Кураре блокирует никотиновые ацетилхолиновые рецепторы в нервно-мышечных синапсах, парализуя двигательные функции и дыхание, хотя сознание и болевая чувствительность сохранны. Курареподобные соединения применяют при хирургических операциях для расслабления мышц.
а) Возбуждающие синапсы. При химической синаптической передаче происходит деполяризация пресинаптического нервного окончания. Высвобождается медиатор (посредник), который связывается с рецепторами мембраны постсинаптической клетки, и открываются ионные каналы.
1. Определение синапса. Нервные клетки передают информацию с помощью потенциалов действия. Переход нервных импульсов с одной клетки на другую осуществляется в морфологически специализированных контактах, синапсах. Поскольку в большинстве случаев, за исключением особого типа ткани — синцития, как плазматические мембраны, так и внутренняя среда смежных клеток лишены непосредственной связи, нервный импульс не может сразу электрически перейти через синапс.
В передаче импульса участвуют особые промежуточные механизмы, такие как высвобождение вещества-посредника (медиатора) в химических синапсах и особое распространение тока в электрических синапсах.
2. Химические синапсы привлекают интерес, в том числе с медицинской точки зрения. В процессе сложных межклеточных взаимодействий здесь могут возникать особые патологические состояния и проявляется чувствительность к определенным фармакологическим препаратам. Таким образом, химические синапсы необходимо рассмотреть достаточно подробно.
3. Структура химических синапсов. Схема химического синапса представлена на рис. 1. Потенциал действия вызывает деполяризацию пресинаптического окончания аксона. В нем содержатся синаптические пузырьки (везикулы), заполненные тысячами молекул медиатора, в данном случае ацетилхолина (АцХ). При деполяризации пресинаптической мембраны с ней сливаются достигшие активных зон везикулы, так что их содержимое высвобождается в синаптическую щель.
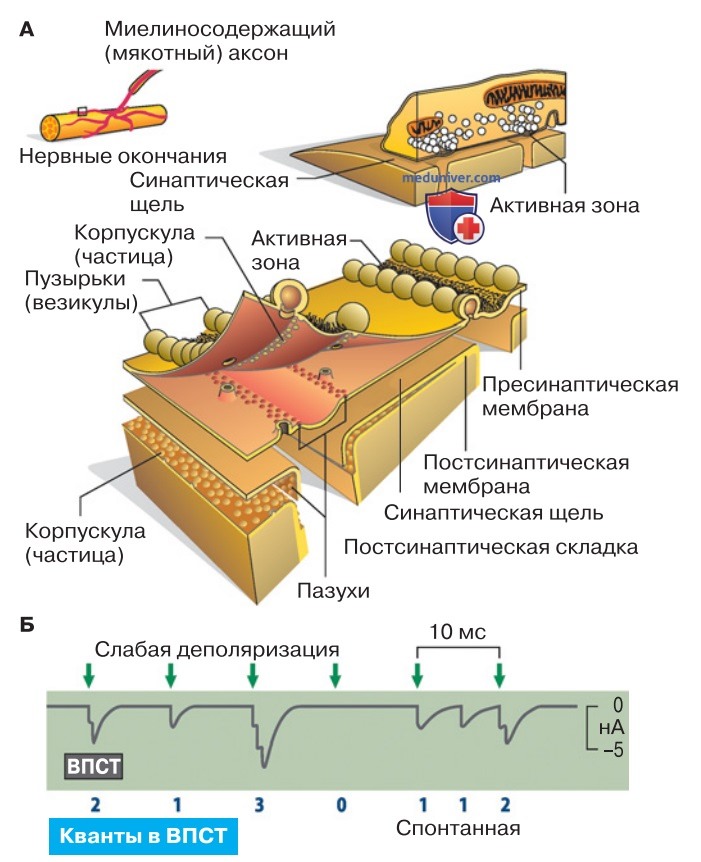
Медиатор диффундирует к постсинаптической мембране, где расположены специфические рецепторы («частицы», рис. 1). При связывании медиатора с рецепторами открываются трансмембранные каналы. Через них проходят ионные токи, благодаря которым изменяется мембранный потенциал постсинаптической клетки; при деполяризации клетки до уровня порога возбуждения генерируется потенциал действия (нервный импульс).
Слабая деполяризация пресинаптического нервного окончания, как правило, способствует высвобождению «квантов» медиатора лишь из небольшого числа везикул; постсинаптическими коррелятами являются квантовые токи концевой пластинки (ТКП) (рис. 1). В нервно-мышечном синапсе (двигательной концевой пластинке) пре-синаптический потенциал действия может инициировать высвобождение медиатора из нескольких сотен везикул.
При этом ток концевой пластинки (рис. 2) будет состоять из суммы сотен квантовых токов и при мембранном потенциале -90 мВ достигать амплитуду 2 нА.
P.S. Семейная гемиплегическая мигрень:
- Симптомы. Семейная гемиплегическая мигрень — это наследственное неврологическое заболевание. Часто оно проявляется односторонней мышечной слабостью или односторонним парезом мышц, вплоть до полного паралича (двустороннего паралича — гемиплегии) и потери сознания. Головная боль у страдающих этим заболеванием может сочетаться с нарушениями равновесия. Последнее обычно связано с повреждением мозжечка, где действительно обнаружены дегенеративные изменения.
- Патогенез. Происходит мутация гена α1-субъединицы потенциалзависимых Са2+-каналов типа P/Q (Cav 2.1), которые характерны для активных зон пресинаптических окончаний (рис. 1), а также присутствуют в больших количествах в клетках Пуркинье мозжечка.
- Лечение. По свидетельствам пациентов, их состояние улучшает верапамил — блокатор Са2+-ка-налов L-типа.
б) Двигательная концевая пластинка. В нервно-мышечном синапсе (двигательной концевой пластинке*) при возбуждении мотонейрона высвобождается ацетилхолин (АцХ). При этом в участке мышечной мембраны, расположенном под пресинаптическим окончанием, открываются катионные каналы и генерируется потенциал концевой пластинки.
P.S. * Существуют различия в определении термина «двигательная концевая пластинка». Есть три варианта определения: а) окончание двигательного аксона, контактирующее с мембраной мышечного волокна, — пресинаптическое окончание; б) область контакта пресинаптического окончания с участком мембраны мышечного волокна, где находятся рецепторы медиатора, — постсинаптическая (или субсинаптическая) мембрана; в) нервно-мышечный синапс, т. е. совокупность (а) и (б).
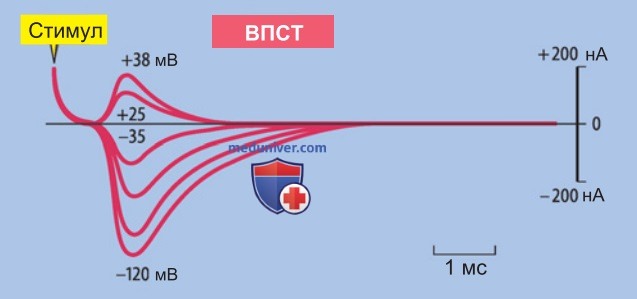
1. Потенциал концевой пластинки. Окончания двигательных аксонов на мышечных волокнах, различимые под лупой, получили название двигательных концевых пластинок. Ток концевой пластинки (ТКП) (в более общем смысле — возбуждающий постсинаптический ток, ВПСТ)** создает местную деполяризацию клеточной мембраны — потенциал концевой пластинки (ПКП) (по смыслу близко к возбуждающему постсинаптическому потенциалу, ВПСП), который может достигать 60 мВ, превышая порог возбуждения и вызывая потенциал действия.
Это означает, что в данном синапсе произошла передача возбуждения от двигательного аксона к мышечному волокну. ТКП протекает по мышечному волокну только в пределах концевой пластинки конкретного мышечного волокна. Возникший там ПКП нарастает до максимума и электротонически распространяется по мышечному волокну, постепенно снижаясь. Все синаптические потенциалы и токи являются по своей природе местными процессами.
P.S. ** Нервно-мышечный синапс относится к возбуждающим синапсам. Исходя из этого автор использовал в статье о нервно-мышечном синапсе наряду с понятиями «потенциал концевой пластинки (ПКП)», «ток концевой пластинки (ТКП)» термины более общего смысла — «возбуждающий постсинаптический потенциал, ВПСП», «возбуждающий постсинаптический ток, ВПСТ».
2. Ток концевой пластинки. Чтобы экспериментально определить зависимость ТКП от электрического напряжения, фиксировали мембранный потенциал на уровне от -120 мВ до +38 мВ (рис. 2). Примерно при -10 мВ направление ТКП изменялось на противоположное. Варьируя концентрации ионов в среде, можно определить, что ТКП возникает вследствие неспецифического повышения мембранной проводимости для ионов Na+, Ca2+ и K+, так что равновесный потенциал соответствует примерно -10 мВ.
Ток концевой пластинки гораздо короче, чем потенциал концевой пластинки (сравните продолжительность ПКП и ТКП на рис. 3). ТКП завершается в течение нескольких миллисекунд, тогда как амплитуда ПКП медленно нарастает по мере того, как падает разряд емкости мембраны, и медленно снижается в зависимости от постоянной времени мембраны τ.
3. Синаптический медиатор. В нервно-мышечном синапсе медиатором (трансмиттером) служит ацетилхолин (АцХ). При местной аппликации АцХ наблюдается деполяризация концевой пластинки. Чувствительность к АцХ ограничена непосредственно областью под нервным окончанием.
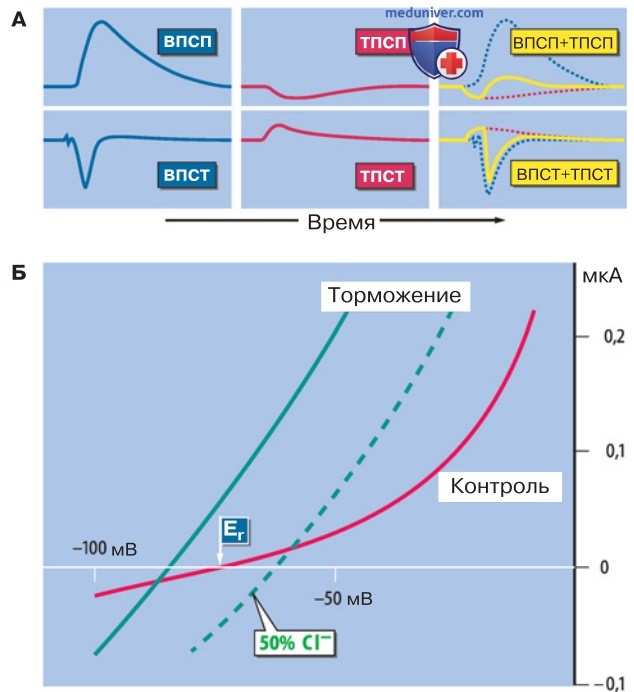
в) Тормозные синапсы. Активация тормозных синапсов ослабляет либо блокирует возбуждение постсинаптической клетки. В организме наряду с возбуждающими синапсами почти столь же часто встречаются синапсы, опосредующие передачу торможения (рис. 3). Слева на рисунке мы видим возбуждающий постсинаптический потенциал (ВПСП) и возбуждающий постсинаптический ток (ВПСТ).
Когда по тормозному аксону поступает потенциал действия, механизм его влияния на постсинаптическую клетку такой же, как в результате поступления импульса от возбуждающего аксона. В постсинаптической клетке развивается тормозный постсинаптический потенциал (ТПСП; обычно это гиперполяризация небольшой амплитуды), которому соответствует выходящий ток. Если возбуждение и торможение инициируются одновременно, то ВПСТ и ТПСТ суммируются.
Однако в итоге сдвиг потенциала получается гораздо меньше, чем сумма раздельных слагаемых (ВПСП + ТПСП), поскольку торможение значительно снижает деполяризацию, вызванную ВПСП, тем самым ослабляя либо блокируя передачу возбуждения в синапсе.
г) Тормозные ионные токи. В тормозных синапсах медиатор открывает каналы для ионов К+ или Cl-. Вследствие этого сопротивление мембраны снижается без существенного сдвига мембранного потенциала, препятствуя деполяризующему влиянию возбуждения.
1. Идентификация ионных токов. Ионные токи во время торможения можно идентифицировать, экспериментируя со сдвигами мембранного потенциала. На рис. 3 представлены графики зависимости тока от мембранного потенциала (вольт-амперные характеристики). «Контрольная» кривая отражает ток фиксации, который требуется (при отсутствии исходного возбуждения мембраны), чтобы деполяризовать либо гиперполяризовать мембрану от уровня потенциала покоя Еr.
При воздействии на клетку тормозного медиатора ГАМК подъем кривой («(+) торможение») становится круче. Теперь кривая пересекает нулевую линию при уровне потенциала на 10 мВ отрицательнее, чем Еr. Это обусловлено гиперполяризацией, ассоциированной с торможением. Чтобы выяснить природу ионных токов при торможении, следует определить равновесные потенциалы для определенных ионов, изменяя их концентрации. Изменение концентрации Na+ или K+ не сопровождалось никакими эффектами.
В то же время при снижении в 2 раза концентрации Cl- прерывистая линия (рис. 3) сместилась вправо почти на 20 мВ, что можно было ожидать исходя из уравнения Нернста. Следовательно, в качестве механизма торможения идентифицировано повышение проводимости мембраны ионов Cl-.
Наиболее распространенный в ЦНС человека тормозный медиатор ГАМК открывает трансмембранные Cl--каналы (ГАМКA-рецепторы) либо К+-каналы (ГАМКB-рецепторы). Другие тормозные медиаторы, например АцХ в синусовом узле сердца, тоже могут открывать К+-каналы. Итак, торможение осуществляется посредством повышения проводимости ионов К+ или Cl-, вследствие которого мембранный потенциал стабилизируется примерно на уровне потенциала покоя.
2. При торможении снижено сопротивление мембраны. Повышение Cl--проводимости во время торможения показано на рис. 3 в виде более крутого подъема кривой ток-потенциал, что соответствует снижению сопротивления мембраны. Пример конкуренции между торможением и возбуждающим током (ВПСТ равен 0,1 мкА) представлен на рис. 3. В контроле ток силой 0,1 мкА деполяризует мембрану от потенциала -74 мВ до -24 мВ, т. е. примерно на 50 мВ, тогда как при конкуренции с торможением наблюдается деполяризация от -84 мВ до -66 мВ, т. е. лишь на 18 мВ.
При снижении сопротивления происходит короткое замыкание возбуждающих токов и возбуждение подавляется. В итоге развивается гиперполяризация.
1. Синапсы. Синапсы — морфологически специализированные контакты, обеспечивающие передачу информации между двумя клетками. Различают две основные формы синапсов: химические синапсы с высвобождением медиатора — вещества-посредника и электрические синапсы с особым распространением тока между контактирующими клетками.
2. Химические синапсы. При деполяризации пресинаптического нервного окончания потенциалом действия (нервным импульсом) высвобождается медиатор, взаимодействующий с рецепторами постсинаптической мембраны. В случае возбуждающей передачи реакция между медиатором и рецептором обеспечивает открывание неспецифических катионных каналов, что приводит к деполяризации постсинаптической клетки/ постсинаптической структуры.
Для нервно-мышечного синапса (двигательной концевой пластинки) свойственно высвобождение в синаптическую щель ацетилхолина. Ацетилхолин вызывает потенциал концевой пластинки, который опосредован током концевой пластинки. Амплитуда потенциала концевой пластинки обычно надпороговая, в отличие от одиночных ВПСП в нейронах.
В тормозных синапсах высвобождение медиатора из пресинаптических нервных окончаний и его взаимодействие с постсинаптическими рецепторами ведет к открыванию К+-каналов и/или Cl--каналов. При этом снижается сопротивление мембраны, а выходящий ионный ток обычно вызывает низкоамплитудную гиперполяризацию (ТПСП). В результате снижается возбудимость клетки: за счет ТПСП мембранный потенциал удаляется от уровня порога, а вследствие пониженного сопротивления происходит короткое замыкание деполяризующих токов.
В итоге мембранный потенциал стабилизируется на уровне потенциала покоя, причем короткое замыкание представляется особенно важным механизмом торможения.
Видео физиология синапса и нерва - профессор, д.м.н. П.Е. Умрюхин
- Рекомендуем ознакомиться далее "Синаптические медиаторы - с точки зрения физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 9.10.2024