
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Синаптические медиаторы - с точки зрения физиологии человека
Оглавление статьи:- Классические медиаторы
- Пептиды и комедиаторы
- Агонисты и антагонисты
- Продолжительность и завершение действия медиаторов
- Коротко
- Список использованной литературы
а) Классические медиаторы. Большинство синаптических медиаторов — низкомолекулярные соединения, например ацетилхолин, ГАМК или глутамат.
1. Низкомолекулярные аминокислоты. Выше упоминались такие медиаторы, как ацетилхолин (АцХ) и ГАМК. Однако медиаторных веществ очень много. Особенно значимые и известные представлены на рис. 4 (вверху). Гамма-аминомасляная кислота (ГАМК) — самый распространенный тормозный медиатор в ЦНС, а более простая по своей структуре аминокислота глицин опосредует торможение мотонейронов.
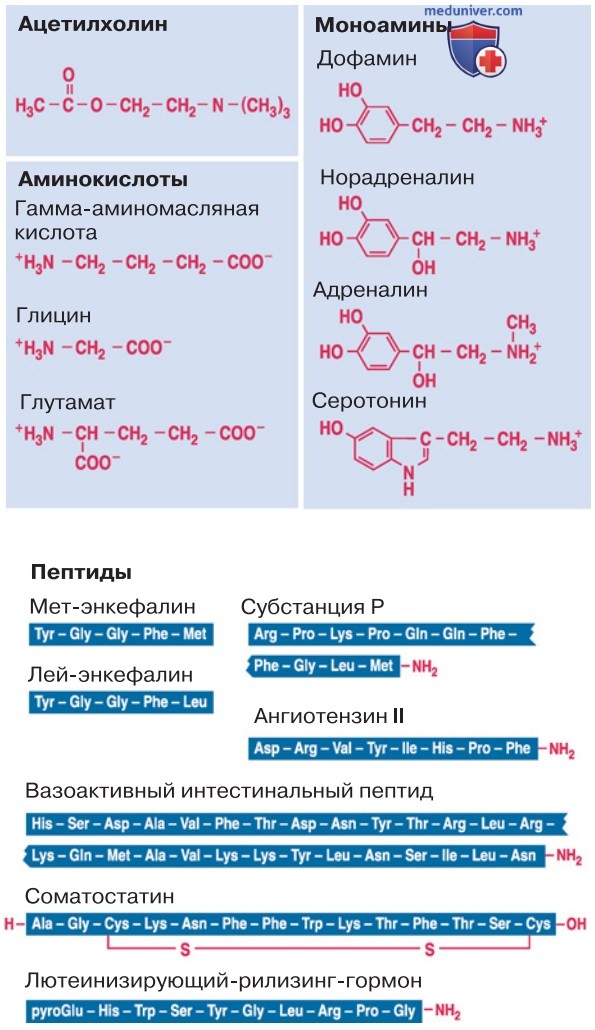
Вверху: классические медиаторы — ацетилхолин, аминокислоты, моноамины. Внизу: пептиды.
Глутамат, который относится к кислым аминокислотам, служит наиболее известным возбуждающим медиатором в ЦНС.
2. Моноамины. Адреналин, норадреналин и дофамин образуют семейство катехоламинов. Эти медиаторы осуществляют возбуждение либо торможение в центральной и периферической нервных системах. Сходное влияние оказывает серотонин (5-гидрокситриптамин, 5-НТ), который вместе с катехоламинами входит в группу моноаминов. К той же группе относится гистамин, возможно, выполняющий медиаторные функции в головном мозге, а также в желудочно-кишечном тракте; однако будучи тканевым гормоном, он в основном опосредует воспалительные реакции.
Все эти «классические» медиаторы представляют собой малые молекулы, зачастую участвующие в межуточном (тканевом) обмене. Когда они связываются со специфическими рецепторами постсинаптической мембраны, повышается проводимость ионных каналов. Если повышение проводимости относится к ионам Na+, Ca2+ и К+, то в синапсе происходит передача возбуждения, если же возрастает проводимость ионов К+ или Cl-, то развивается торможение.
б) Пептиды и комедиаторы. Пептиды обладают относительно медленным синаптическим эффектом; часто они сосуществуют с классическими медиаторами.
1. Пептидные медиаторы. Наряду с классическими медиаторами на рис. 4 (внизу) представлен ряд пептидных медиаторов. Эти вещества обнаружены в ЦНС и/или вегетативной нервной системе, причем механизм их действия не вполне выяснен. Часто они являются синаптическими модуляторами: не влияют непосредственно на проводимость синаптических мембран, но изменяют интенсивность и длительность действия классических медиаторов, причем иногда высвобождаются вместе с другими медиаторными веществами.
На рис. 4 представлены наиболее значимые пептиды.
- Энкефалины связываются с опиоидными (опиатными) рецепторами, взаимодействующими с морфином, и наряду с прочими функциями опосредуют болевую чувствительность.
- Вещество Р тоже имеет отношение к болевой чувствительности, а помимо этого вызывает сокращение гладкой мускулатуры.
- Ангиотензин II — гормон, который оказывает сильное влияние на кровеносные сосуды, а также действует на нейроны ЦНС.
- Вазоактивный интестинальный полипептид (ВИП), соматостатин и люлиберин (LHRH, рилизинг-фактор лютеотропина — лютеинизирующего гормона) участвуют в регуляции выработки гормонов в гипофизе, а также активны в синапсах.
P.S. Сомедиаторы (комедиаторы). Долгое время полагали, что нервная клетка высвобождает из окончаний своего аксона единственный медиатор (принцип Дейла). Однако показано, что из клеток вегетативной нервной системы, по крайней мере на этапе эмбрионального развития, высвобождается не только АцХ, но и адреналин. В нервно-мышечных синапсах и в вегетативной нервной системе совместно с АцХ и катехоламинами высвобождается также аденозинтрифосфат (АТФ), который в результате дефосфорилирования преобразуется в аденозин и становится медиатором.
Нередко вместе с классическим медиатором, таким как норадреналин, из пресинаптических окончаний выделяется пептид, участвующий в синаптической передаче. Особенности взаимодействия медиаторных веществ и сомедиаторов до конца не выяснены, чаще всего их трактуют как модуляцию.
Сведения о медиаторах, способных к трансклеточной диффузии, например NO.
в) Агонисты и антагонисты. Агонисты — это вещества, взаимодействие которых с синаптическими рецепторами вызывает такой же эффект, как медиаторы, тогда как антагонисты блокируют влияние медиаторов.
1. Агонисты. В результате взаимодействия рецепторов постсинаптической мембраны со специфическим для них медиатором повышается проводимость определенных ионов. Тем не менее специфичность медиатора не абсолютна, практически все рецепторы способны связываться и с другими веществами. Если связывание вещества с рецептором приводит к свойственному для него, биологически ожидаемому изменению проводимости, то это вещество полностью заменяет медиатор и называется агонистом.
Для рецепторов концевой пластинки агонистами служат, например, карбамилхолин и суберилдихолин. Другие соединения тоже связываются с рецепторами, но они малоэффективны применительно к сдвигу ионной проводимости. Это частичные агонисты, например холин для концевой пластинки.
2. Антагонисты. Существуют также вещества, которые связываются с синаптическими рецепторами, но не изменяют проводимость. Оккупируя рецепторы, они препятствуют действию агонистов. Такие вещества называются антагонистами. Если антагонист конкурирует с агонистами за участок связывания, то это конкурентный антагонист. Если же вещества мешают действию агонистов без конкуренции за участок связывания, то это неконкурентные антагонисты.
3. Расслабление мышц. Известным конкурентным антагонистом влияния АцХ на двигательную концевую пластинку является кураре (d-тубокурарин) — яд, которым индейцы смазывали стрелы. По мере повышения концентрации кураре возрастает количество заблокированных рецепторов и постепенно ослабевает эффект от связывания АцХ с рецепторами, которые остались интактными. Под влиянием кураре падает амплитуда потенциала концевой пластинки (рис. 5); при достаточно высокой дозе уже невозможно достигнуть порога генерации потенциала действия: мышца парализована.
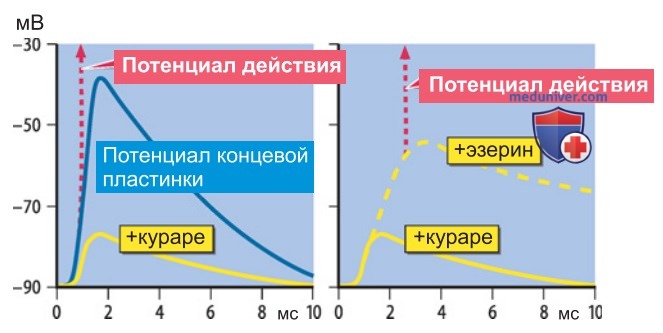
Курареподобные вещества применяются при анестезии для расслабления мышц. При полном расслаблении необходимо поддерживать дыхание пациента искусственным путем. Мышечное расслабление иного типа вызывает агонист длительного действия, такой как сукцинилхолин, обусловливающий продолжительную деполяризацию концевой пластинки. Деполяризация инактивирует Na'-ка-налы мышечной мембраны, препятствуя возбуждению мышцы.
P.S. Фармакологические аспекты. Агонисты и антагонисты часто используют в физиологии для выяснения механизмов синаптической передачи, а также в клинических целях при поиске подходов к лечению. Вместе с тем взаимодействие агонистов и антагонистов составляет предмет фармакологического анализа. Изучая эффективность различных агонистов и антагонистов, можно также классифицировать различные типы рецепторов определенных медиаторов, например рецепторов АцХ или адреналина.
г) Продолжительность и завершение действия медиаторов. Эффекты медиаторов завершаются либо благодаря расщепляющим ферментам (например, холинэстеразе), либо путем активного захвата соседними клетками, либо в результате дальнейшей диффузии.
1. Продолжительность действия. При диффузии медиатора из узкой синаптической щели (рис. ниже) его концентрация должна снижаться достаточно долго. Однако эффекты большинства медиаторов имеют короткие сроки, максимум до тех пор, пока продолжается постсинаптический ток. Таким образом, действие медиаторов ограничено во времени. Это обусловлено двумя основными механизмами: расщеплением и обратным захватом медиатора.
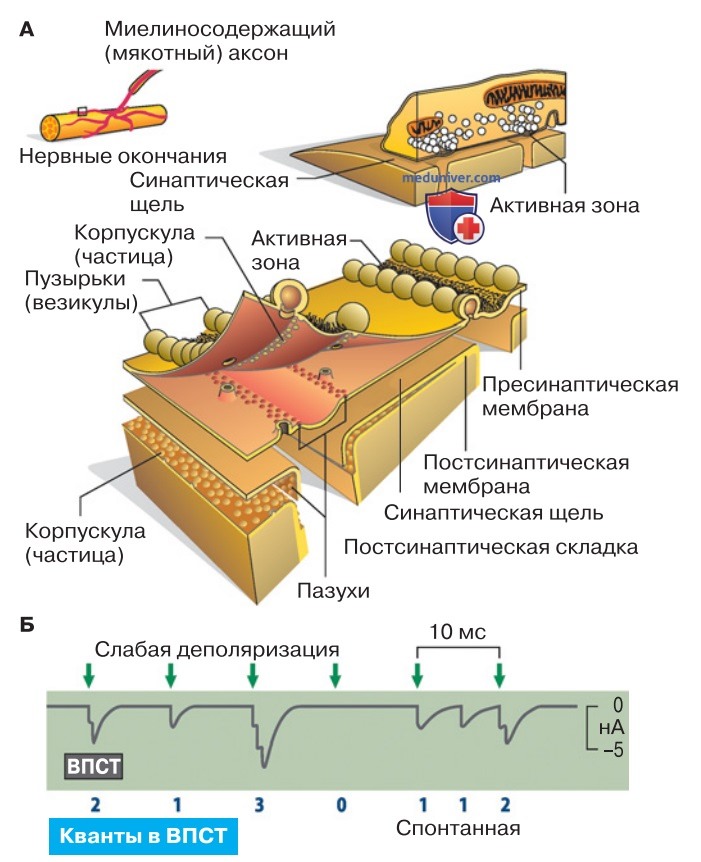
2. Расщепление медиатора ферментом. Двигательная концевая пластинка обладает очень эффективной системой расщепления АцХ. На постсинаптической мембране присутствует в высокой концентрации ацетилхолинэстераза — фермент, расщепляющий АцХ на ацетат и холин (рис. выше). Значительная часть АцХ, высвобождаемого в синаптическую щель, расщепляется прежде, чем он достигнет рецепторов; ацетилхолинэстераза разрушает практически весь АцХ в среднем менее чем за 0,1 мс. Таким образом синапс быстро восстанавливается для следующего цикла передачи сигнала.
P.S. Миастения:
- Симптомы. Миастения (myasthenia gravis) — аутоиммунное заболевание с поражением нервно-мышечных синапсов. Пациентам свойственны повышенная утомляемость, мышечная слабость, параличи. Начальные признаки заболевания нередко обнаруживаются на глазных мышцах (раздвоение изображений). Характерно, что кураре в небольших дозах (так же как физическое усилие) усугубляет симптомы, а блокаторы ацетилхолин-эстеразы (эзерин или физостигмин) приносят временное улучшение.
- Патогенез. У некоторых пациентов обнаруживаются опухоли тимуса (вилочковой железы), удаление которых влияет позитивно. У экспериментальных животных, иммунизированных никотиновыми ацетилхолиновыми рецепторами, проявлялись типичные симптомы миастении. Введение здоровым животным антител к соответствующим рецепторам сопровождалось развитием типичных симптомов миастении. Антитела к никотиновым ацетилхолиновым рецепторам выявляются у многих больных миастенией.
Антитела вызывают локальные повреждения структуры рецепторов, что нарушает их функции, стимулируя дальнейшую дезорганизацию. Аутоантитела к ионным каналам глутаматерги-ческих рецепторов обнаружены при редкой форме эпилепсии, энцефалите Расмуссена.
- Лечение. Наряду с блокаторами ацетилхолин-эстеразы (неостигмином или пиридостигмином) при миастении применяются по показаниям иммунодепрессанты (например, глюкокортикоиды).
3. Ацетилхолинэстераза. Роль ацетилхолин-эстеразы (АХЭ) в передаче сигнала через нервно-мышечный синапс доказывают эксперименты со специфическими блокаторами. На рис. 2 продемонстрировано действие одного из них, эзерина (физостигмина): длительность и амплитуда потенциала концевой пластинки (ПКП) возрастают по сравнению с нормой, поскольку АцХ сохраняет высокую концентрацию, дольше воздействуя на рецепторы. В случае, представленном на рис. 2, это «терапевтический эффект», поскольку влияние эзерина исследуется на мышце, парализованной кураре.
Полученный эффект повышения амплитуды ПКП позволяет достигнуть порога возбуждения мышцы, тем самым снимая паралич.
Ингибиторы АХЭ используются для восстановления мышечного тонуса после анестезии, а также при клинической картине генерализованной миастении. Вместе с тем ингибиторы АХЭ часто применяются в качестве инсектицидов и могут стать причиной отравления. Некоторые боевые отравляющие вещества блокируют АХЭ; затягивая процесс холинергической синаптической передачи, они вызывают пароксизмальные вегетативно-сосудистые проявления.
4. Обратный захват медиатора. Во многих синапсах медиатор удаляется из синаптической щели посредством транспортных механизмов в мембранах примыкающих клеток. Механизмы захвата особенно важны для адреналина, норадреналина, ГАМК и глутамата. В холинергических синапсах транспортируется не АцХ, а продукт его распада, холин. Такого рода обратный транспорт в нервное окончание ограничивает потребности в ресинтезе медиатора.
Наряду с расщепляющими ферментами механизм обратного захвата медиаторов составляет основу влияния фармакологических средств на синаптическую передачу.
5. Диффузия медиатора. После высвобождения медиатор диффундирует из синаптической области с постоянной времени ~100 мкс. Диффузия способствует быстрому завершению синаптической передачи. Наличие дополнительных механизмов распада и обратного транспорта медиатора еще раз указывает, насколько важен контроль его концентрации.
1. Синаптические медиаторы. К классическим медиаторам (нейромедиаторам) относятся АцХ, ГАМК, глицин, глутамат, дофамин, норадреналин, адреналин, серотонин и другие низкомолекулярные соединения.
Существуют также пептидные медиаторы, например энкефалины и вещество Р, которые в качестве синаптических модуляторов опосредуют достаточно медленные синаптические эффекты. Они влияют на интенсивность и продолжительность действия классических медиаторов и часто содержатся в пресинаптических окончаниях совместно с классическими медиаторами.
Рецепторы медиаторных веществ не обладают абсолютной специфичностью. Практически для любого рецептора имеются вещества, которые связываются с ним и могут служить лекарственными средствами. Это вещества двух типов:
• агонисты — действуют подобно медиаторам;
• антагонисты — блокируют действие медиаторов.
2. Расщепление и обратный захват медиаторов. Время воздействия медиаторов на рецепторы ограничивают расщепляющие ферменты (например, ацетилхолинэстераза двигательной концевой пластинки), активное поглощение пресинаптическим нервным окончанием (обратный захват медиатора), а также соседними глиальными клетками и, наконец, диффузия в интерстициальное пространство.
Видео строение синапса
- Рекомендуем ознакомиться далее "Взаимодействие синапсов - с точки зрения физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 9.10.2024