
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Синаптическая пластичность - с точки зрения физиологии человека
Оглавление статьи:- Долговременная потенциация
- Долговременная депрессия
- Динамическая регуляция и пластичность синапсов
- Коротко
- Список использованной литературы
а) Долговременная потенциация. Долговременная потенциация соответствует продолжительному повышению эффективности синаптической передачи.
1. Возможные механизмы научения. Основополагающую способность даже примитивных нервных систем составляет научение, т. е. изменение реакции нервной системы на основе опыта. Более подробно о научении пойдет речь в отдельной статье на сайте (просим Вас пользоваться формой поиска по сайту выше).
Здесь же будут рассмотрены два типа синаптических реакций — долговременная потенциация (ДВП) и долговременная депрессия (ДВД), которые обсуждаются в качестве возможных клеточных механизмов научения.
ДВП исследована, в частности, в пирамидных клетках гиппокампа. Для ее развития требуется мощная активация синаптического входа этих клеток высокочастотной серией импульсов, после которой синаптическая передача с этого входа может продолжаться целыми днями.
2. Долговременная потенциация. ДВП выявлена в глутаматергических синапсах. Например, на дендритных шипиках пирамидных клеток находятся глутаматергические рецепторы NMDA-типа, а также АМРА/каинатного типа (А/К-типа), т. е. не-NMDA-типа.
Одиночный импульс от афферентного нерва вызывает высвобождение глутамата в достаточном количестве для открывания каналов А/К-типа (рис. 1). Что касается каналов NMDA-типа, то они не открываются, потому что блокированы ионами Mg2+ (рис. ниже). В ответ на такой импульс возникает ВПСП сравнительно небольшой амплитуды.
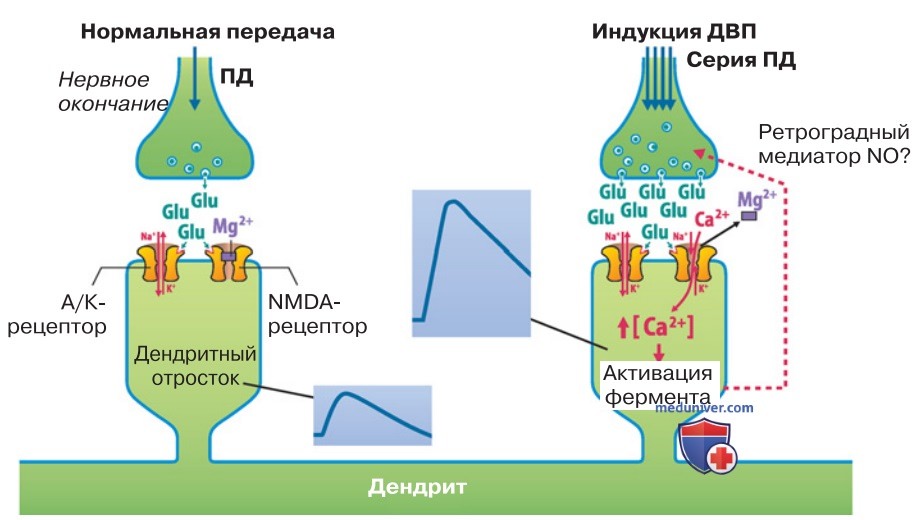
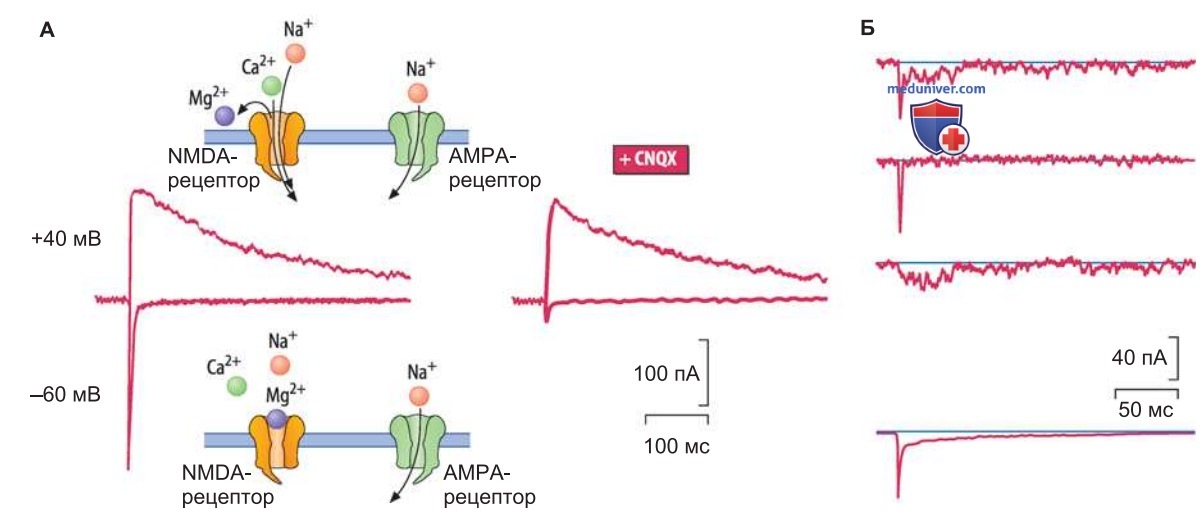
Когда от афферентных аксонов поступает более продолжительная серия импульсов (рис. 1), концентрация глутамата в шипике сильно возрастает и открывается большее число каналов А/К-типа. В результате деполяризация мембраны пирамидной клетки достигает достаточного уровня для того, чтобы блокада потенциалзависимых NМDА-каналов прекратилась и произошла генерация высокоамплитудного ВПСП.
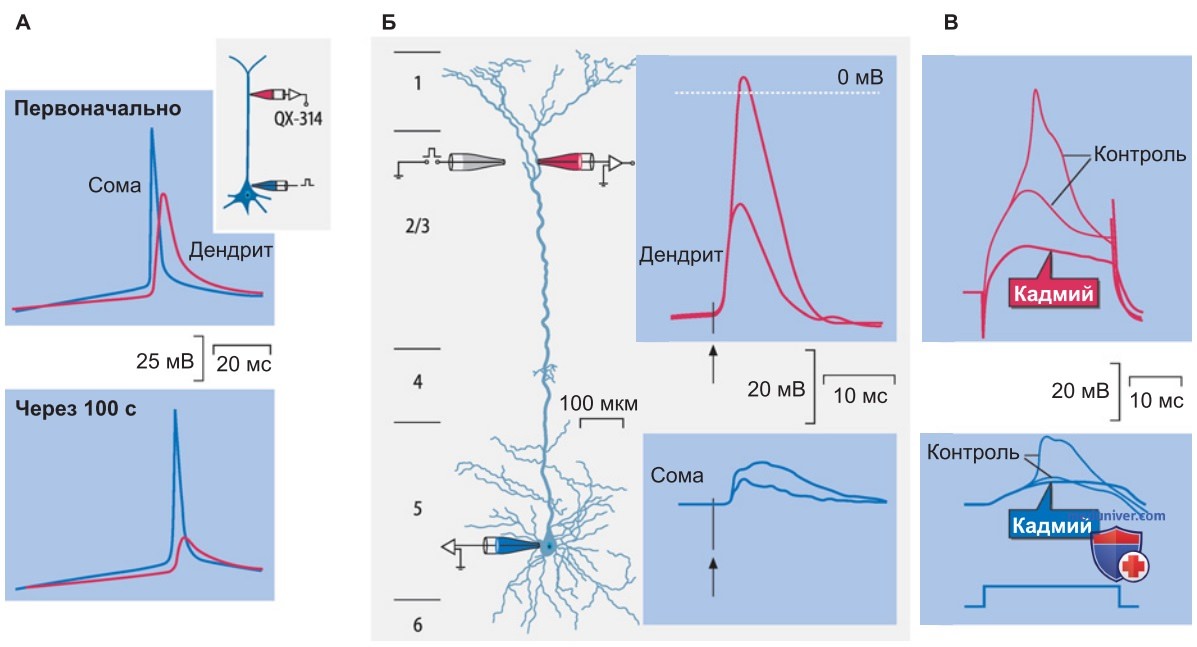
Деполяризацию, необходимую для открывания NМDА-каналов, могут также вызвать ретроградные потенциалы действия (рис. ниже).
Важно отметить, что при открывании NMDA-каналов в клетку должно поступить достаточное количество Са2+ для активации различных ферментных систем. В итоге вследствие встраивания рецепторов возрастает постсинаптическая чувствительность к глутамату и/или вследствие активации NO-синтазы образуется NO, который диффундирует к пресинаптическим окончаниям и усиливает высвобождение медиатора. Наряду с постсинаптическими механизмами обнаружены и пресинаптические механизмы ДВП.
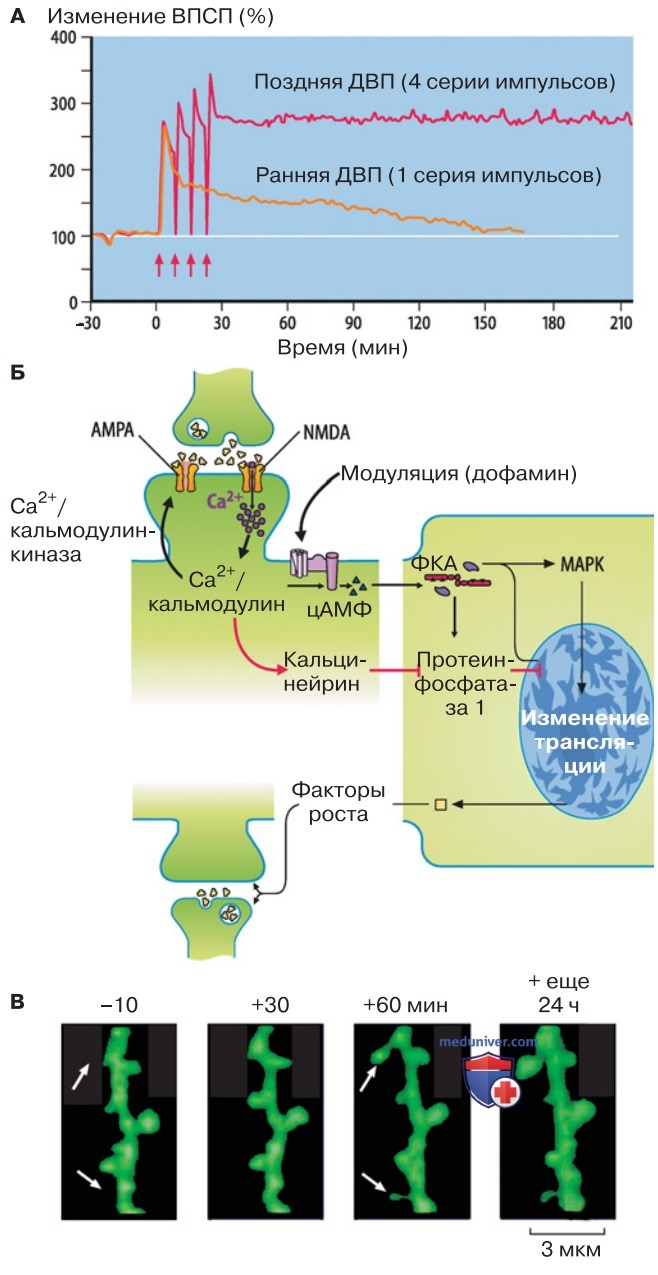
ДВП может продолжаться в течение 1-2 ч и гораздо дольше (рис. 2). Позднюю фазу ДВП можно прервать блокадой синтеза белка, следовательно, на этом этапе вовлечена транскрипция генов (рис. 2). Возможны морфологические изменения — увеличение размера и числа синапсов, что указывает на реорганизацию дендритных ши-пиков (рис. 2).
б) Долговременная депрессия. Долговременная депрессия соответствует продолжительному снижению эффективности синаптической передачи.
Долговременная депрессия (ДВД) — явление, по знаку противоположное ДВП, подробно исследованное в клетках Пуркинье, которым принадлежат эфферентные волокна из мозжечка. Эти клетки получают три входа. Когда два из трех входов, лиановидные (лазящие) волокна и параллельные волокна, возбуждаются одновременно, то передача сигналов от параллельных волокон к клеткам Пуркинье на несколько часов блокируется вследствие развития ДВД.
ДВД обусловлена одновременной активацией двух типов ионных каналов глутаматергических рецепторов. Первые из них относятся к классическому типу А/К-рецепторов и активируются при стимуляции лиановидных волокон. Возникает сильная деполяризация с интенсивным входом Са2+ в клетки.
Если одновременно стимулируется другой вход, через параллельные волокна, то вследствие высокой концентрации выделенного глутамата активируются и метаботропные рецепторы. Они сопряжены с G-белком и запускают сигнальный каскад инозитолтрифосфата (IP3), обеспечивающего высвобождение Са2+ из внутриклеточных запасов.
В клетках создается высокая концентрация Са2+, достаточная для активации синтеза NO (аналогично рассмотренной выше ситуации при развитии ДВП). В свою очередь, NO способствует образованию цГМФ; развивается продолжительная десенситизация рецепторов А/К-типа и как следствие — ДВД.
в) Динамическая регуляция и пластичность синапсов. В некоторых синапсах высокочастотные серии импульсов порождают ДВП, а низкочастотные — ДВД.
1. Роль последовательности синаптических событий. Реализация одной либо другой формы долговременной пластичности (ДВП либо ДВД) может зависеть от соотношения между активностью синаптических входов и самой постсинаптической клетки. Ретроградное проведение ПД (рис. выше) осуществляет «контроль результата» даже в дистальных дендритах.
Если слабый вход (который сам по себе не вызывает ПД в постсинаптической клетке) задействован незадолго до сильного входа (вызывающего ПД), то можно ожидать развитие ДВП (рис. 3). В случае обратной последовательности событий итогом активации слабого входа становится ДВД (рис. 3). В иных синапсах эффект (выбор между ДВП и ДВД) может быть противоположным.
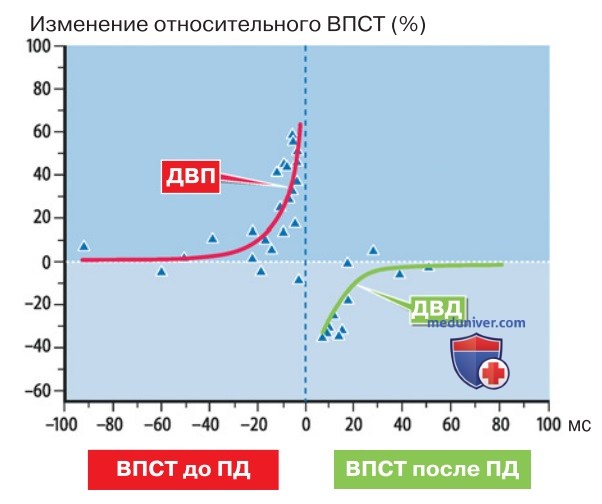
Это наблюдение свидетельствует, насколько существенна последовательность синаптических событий в определении формы пластичности.
2. Пластичность и онтогенез. Степень пластичности синапсов зависит от стадии развития нервной системы. Обычно незрелые синаптические входы более пластичны, чем зрелые. По мере созревания нервной системы количество синапсов уменьшается (рис. 4). Как показано на рисунке, к моменту рождения мыши два аксона образуют окончания на волокне скелетной мышцы. Одно нервное окончание продолжает свой рост в течение нескольких дней, остальные деградируют.
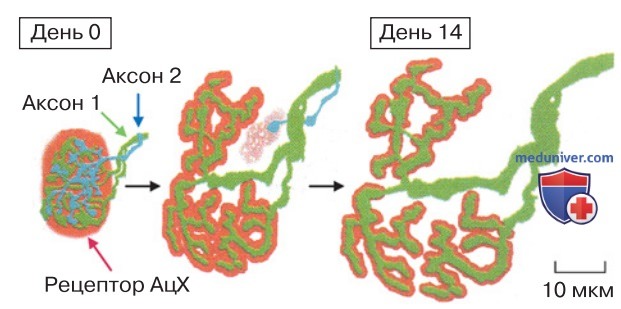
В созревании синапсов важным фактором является также соотношение между активностью синаптических входов и электрическим ответом постсинаптической клетки.
г) Коротко. Синаптическая пластичность. Две формы долгосрочных изменений эффективности синаптической передачи считаются возможными механизмами научения. Долговременная потенциация (ДВП) развивается в случае сильной активации синаптического входа серией высокочастотных потенциалов действия. В такой ситуации повышенная эффективность постсинаптической клетки может сохраняться в течение нескольких дней. Долговременная депрессия (ДВД) означает продолжительное снижение эффективности синаптической передачи.
В некоторых синапсах при сериях высокочастотных импульсов наблюдается ДВП, а при низкочастотных — ДВД. Таким образом, обеспечивая продолжительное усиление или уменьшение эффективности синапсов, ЦНС, ДВП и ДВД могут участвовать в процессе научения.
Видео физиология синапса и нерва - профессор, д.м.н. П.Е. Умрюхин
- Рекомендуем ознакомиться далее "Электрическая синаптическая передача - с точки зрения физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 9.10.2024