
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Взаимодействие синапсов - с точки зрения физиологии человека
Оглавление статьи:- Пространственная и временная суммация
- Потенциалы действия в дендритах
- Постсинаптическое и пресинаптическое торможение
- Гетеросинаптическое облегчение
- Коротко
- Список использованной литературы
а) Пространственная и временная суммация. Ионные токи и потенциалы множества синапсов, находящихся на нервной клетке, суммируются, если они возникают в разных синапсах одновременно или последовательно друг за другом в одном и том же синапсе.
Двигательная концевая пластинка — особенно удобный для исследования тип синапса. Каждое мышечное волокно имеет, как правило, только один синапс; при возбуждении двигательного аксона возникает надпороговый потенциал концевой пластинки, так что каждый потенциал действия, поступивший по двигательному нерву, приводит к мышечному сокращению.
В большинстве синапсов, прежде всего в ЦНС, одиночный синаптический потенциал является, напротив, подпороговым, не превышая 1 мВ. Зато на постсинаптических клетках находится множество возбуждающих синапсов, зачастую несколько тысяч, так что их эффекты суммируются; возбуждению противостоит влияние примерно такого же числа тормозных синапсов. Эти синапсы принадлежат многочисленным нейронам, аксоны которых конвергируют на исследуемой клетке.
1. Пространственная суммация. На рис. 1 представлено только два синапса из тысяч возбуждающих синапсов на одной нервной клетке, чтобы продемонстрировать их взаимодействие. В обоих синапсах в клетке регистрируется непродолжительный ток (возбуждающий постсинаптический ток, ВПСТ); он вызывает местный сдвиг потенциала — возбуждающий постсинаптический потенциал, ВПСП (рис. ниже). Выход токов частично совершается на некотором расстоянии от того и другого синапса, в частности в аксонном холмике, где тело клетки переходит в аксон (рис. 1).
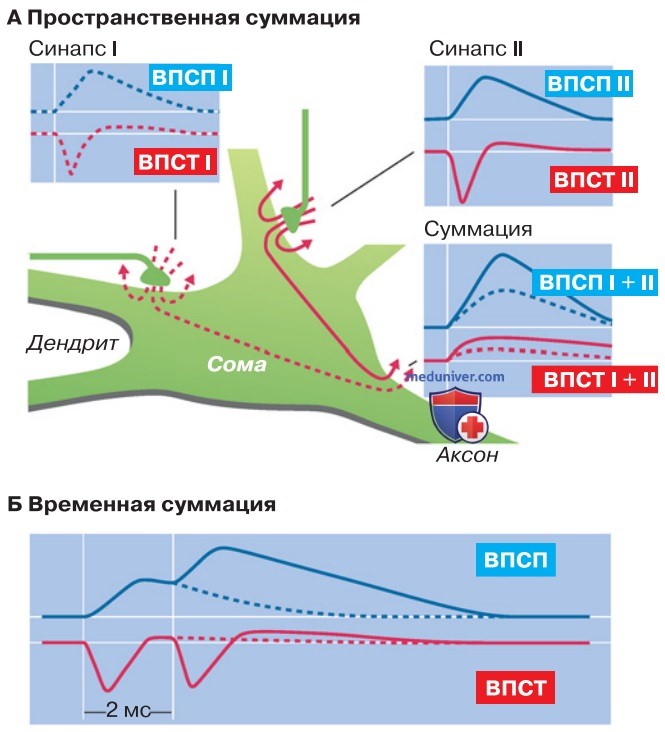
Одиночный ВПСП, электротонически распространяющийся к аксонному холмику, невелик по амплитуде, однако благодаря суммации токов от двух одновременно активированных синапсов получается более значительный ВПСП. Поскольку здесь взаимно добавляются эффекты одновременной активации пространственно разделенных синапсов, данное явление получило название пространственной суммации.
2. Аксонный холмик — место суммации. Электротонически распространяющиеся ВПСП суммируются друг с другом в любой части клетки. Однако начальный сегмент эфферентного аксона не случайно представлен на рис. 1 в качестве места суммации. Дело в том, что у большинства нейронов сома и дендриты невозбудимы или имеют высокий порог возбуждения. Аксон, напротив, достаточно легко возбудим, так что потенциал действия (нервный импульс) генерируется, как правило, в так называемом аксонном холмике — участке, где аксон отходит от тела клетки.
С учетом высокой плотности Na+-каналов на аксонном холмике суммация сдвигов потенциала именно в этом участке определяет, обеспечат ли местные синаптические потенциалы генерацию распространяющегося возбуждения.
3. Временная суммация. Еще одна форма синаптической суммации представлена на рис. 1. Речь идет о ситуации, когда один и тот же синапс или синапсы, находящиеся по соседству друг с другом, активируются в течение короткого промежутка времени (до нескольких миллисекунд). В этом случае синаптические токи практически полностью заканчиваются еще до начала следующего акта возбуждения.
Однако синаптические потенциалы распространяются медленнее, чем токи, так что после завершения синаптического тока емкость мембраны продолжает разряжаться с постоянной времени электротонического потенциала. Если раньше, чем закончится разряд мембраны, возникнет новый синаптический ток, то вызванная им деполяризация добавляется к еще оставшейся. Это явление носит название временной суммации.
В реальной нервной клетке при множестве синапсов и высокой частоте их активации обе формы суммации, временная и пространственная, происходят одновременно, устанавливая изменчивый уровень деполяризации, который определяет частоту генерации потенциалов действия в аксоне.
б) Потенциалы действия в дендритах. Дендриты тоже могут активно реагировать на деполяризацию открытием Na+- или Ca2+-каналов. В результате происходит модификация синаптических потенциалов или же синапсы подвергаются пластическим изменениям.
1. Ретроградное (обратное) распространение потенциала действия. Аксонный холмик рассматривался в качестве места первичного возникновения потенциалов действия (нервных импульсов). Как уже говорилось, от аксонного холмика импульсы активно проводятся далее по аксону. Однако потенциалы действия поступают от аксонного холмика и в обратном направлении: по дендритному дереву происходит ретроградное распространение нервных импульсов.
Дендриты, схематично представленные на рис. 1, могут достигать нескольких миллиметров в длину и интенсивно ветвятся (рис. 2).
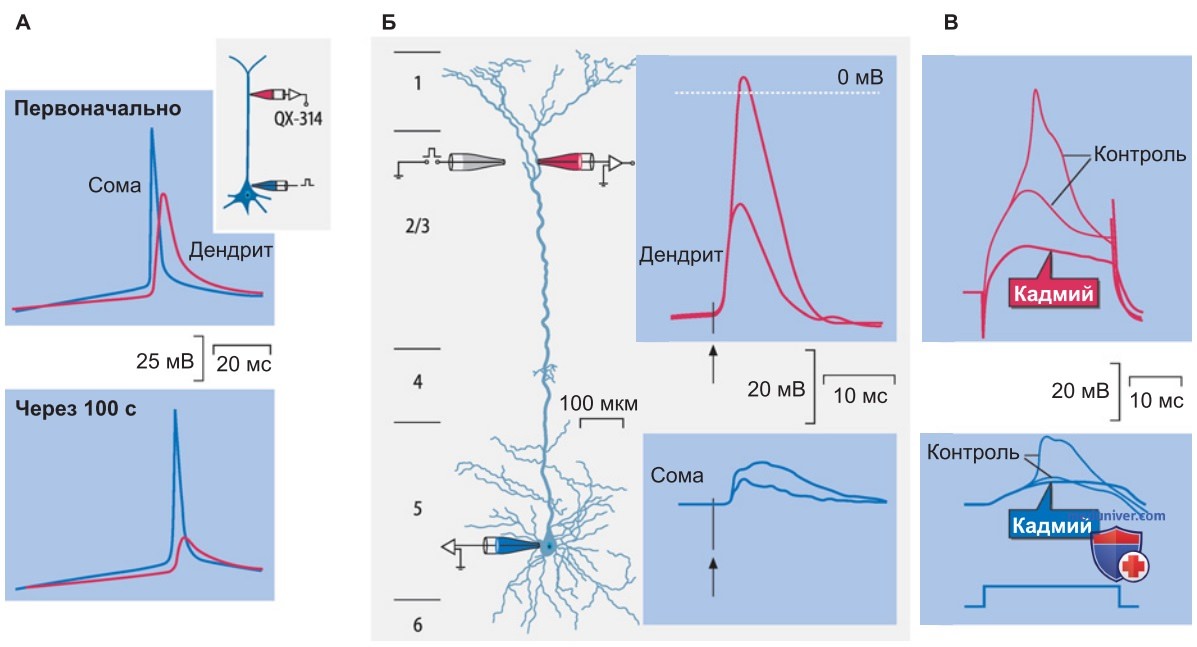
На рис. 2 представлен потенциал действия, вызванный одиночным стимулом в соме пирамидной клетки ЦНС. С небольшой задержкой и несколько сниженный по амплитуде, потенциал действия зарегистрирован и в дендрите. Поскольку в дендритах были блокированы Na+-каналы, дендритный ответ на соматический потенциал действия сильно снизился.
В данном случае дендрит был возбудимым и, так же как аксон, активно проводил потенциалы действия. Ретроградное распространение импульсов в дендритном дереве играет существенную роль в пластических изменениях синапсов.
2. Кальциевые потенциалы действия. В дендритах могут генерироваться потенциалы действия (рис. 2). Надпороговый стимул вызывает в дендрите относительно длительный потенциал действия. В соме нейрона при этом регистрируется медленная подпороговая деполяризация (рис. 2). Если Ca2+-каналы дендритов блокированы ионами кадмия (рис. 2), реакция дендрита на деполяризующий стимул становится практически электротонической, и соответственно сигнал, поступающий к соме клетки, значительно снижен и замедлен по сравнению с контролем, когда этот дендрит был способен генерировать кальциевые потенциалы действия.
Явления, вызванные в эксперименте стимулирующим импульсом тока, характерны и для естественной синаптической активности, например при взаимодействии ВПСП и ретроградно поступивших потенциалов действия. В это время в дендритах может происходить значительное местное повышение концентрации Ca2+, вызывающее изменения их структуры Характер ответов дендрита на деполяризацию зависит от состояния местных Na+-, Ca2+- или K+-каналов. Вариабельность местной экспрессии ионных каналов позволяет нейронам ЦНС адекватно реагировать на различные синаптические сигналы.
в) Постсинаптическое и пресинаптическое торможение. При постсинаптическом и пресинаптическом торможении блокируется возбуждение; во втором случае снижается либо прекращается высвобождение медиатора.
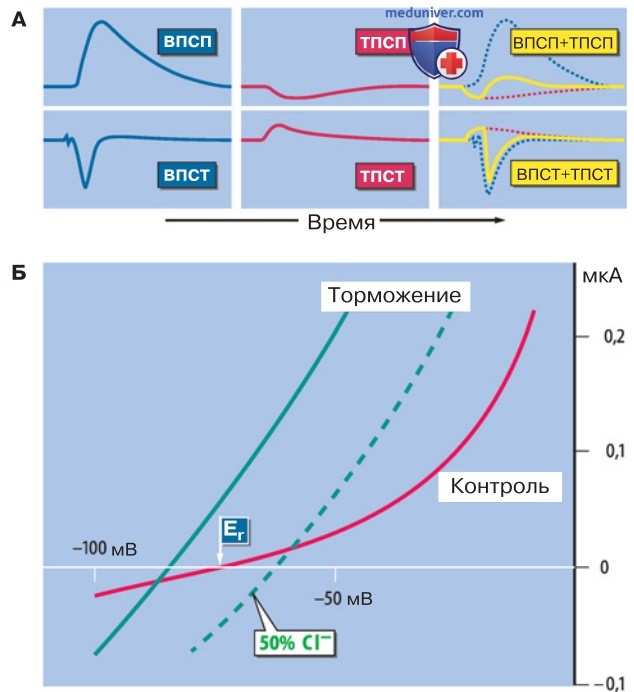
1. Постсинаптическое торможение. Один из видов взаимодействия синапсов в клетке — это постсинаптическое торможение. Как показано на рис. выше, во время торможения шунтируются возбуждающие постсинаптические потенциалы (ВПСП). Тормозные постсинаптические потенциалы (ТПСП) зачастую дополнительно гиперполяризуют мембрану, препятствуя тому, чтобы деполяризация достигла порогового уровня возбуждения.
2. Плотность размещения тормозных синапсов на теле нейрона. В нервной клетке осуществляется временная и пространственная суммация ТПСП и ТПСТ друг с другом, а также с ВПСП. Общий результат суммации множества постсинаптических потенциалов определяет частоту потенциалов действия в аксоне. При этом важную роль играет пространственное распределение возбуждающих и тормозных синапсов. Нередко от тормозных синапсов, плотно размещенных на соме недалеко от выхода аксона, зависит, в какой мере повлияет на аксон деполяризация в результате ВПСП, который возникает преимущественно в дендритах.
3. Пресинаптическое торможение. Эта сложная форма торможения осуществляется при непосредственном контакте возбуждающего синапса с тормозным аксо-аксонным синапсом. Такое торможение можно продемонстрировать на примере мотонейрона (рис. 3). Мотонейроны получают функционально важный возбуждающий синаптический вход от мышечных веретен через посредство афферентных волокон группы Ia. Располагающиеся на мотонейроне окончания Ia-афферентов образуют аксо-аксонные синапсы с окончаниями нервных волокон от интернейронов.
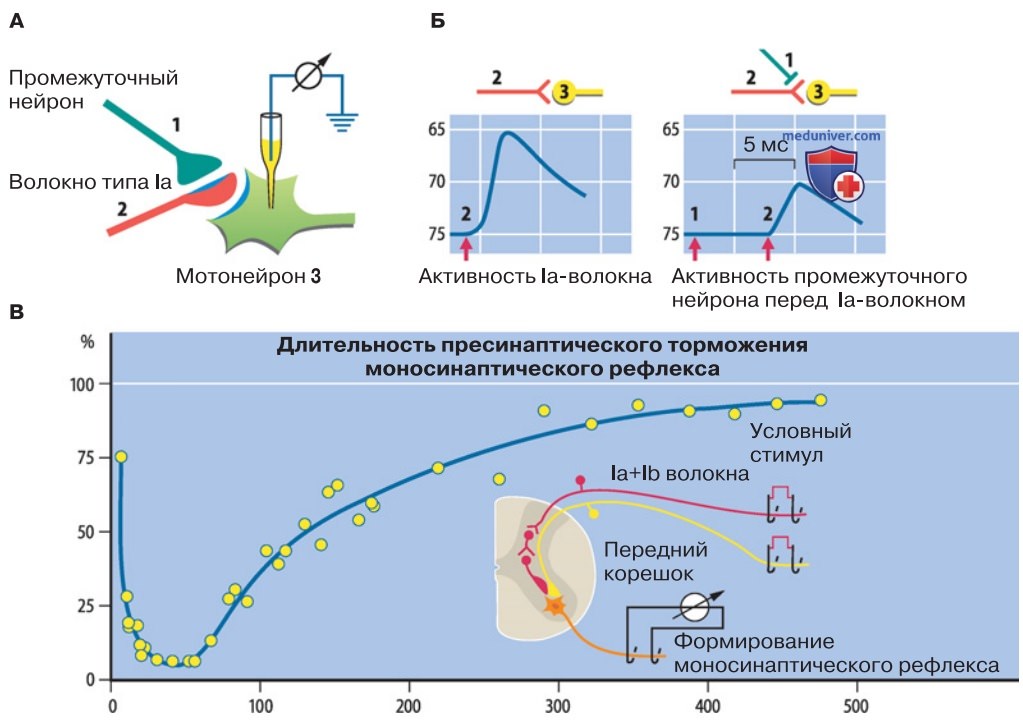
Если возбуждение этих интернейронов произойдет на несколько миллисекунд раньше, чем возбуждение Ia-афферентов, то ВПСП, вызванный Ia-афферентами в мотонейроне, будет заторможен (рис. 3). Как показала регистрация индуцированной пресинаптическим торможением депрессии моносинаптического рефлекса, длительность этого торможения превышала 100 мс (рис. 3).
Пресинаптическое торможение служит эффективным механизмом регуляции двигательных функций спинного мозга. Его особое преимущество — возможность целенаправленного ингибирования отдельных синаптических входов без изменения общей возбудимости клетки. При этом «нежелательную» информацию можно блокировать прежде, чем произойдет интеграция на уровне тела нервной клетки. Функциональная значимость пресинаптического торможения в спинном мозге демонстрируется в эксперименте с блокированием ГАМКергических синапсов антагонистом ГАМК бикукуллином: в мышцах начинаются судороги.
При исследовании механизма торможения нервных окончаний Ia-афферентов была зарегистрирована значительная деполяризация, опосредованная химическими ГАМКергическими синапсами от окончаний интернейронов. Деполяризация первичных афферентов инактивирует возбуждающие Na+-каналы на окончаниях Ia-афферентов, тем самым блокируя генерацию потенциалов действия.
г) Гетеросинаптическое облегчение. При гетеросинаптическом облегчении (фасилитации) усиливаются одновременно два синаптических входа к нейрону.
1. Взаимодействие синаптических входов. Так же как в случае пресинаптического торможения, взаимодействие двух разных синаптических входов к одному сложному синапсу усиливает возбуждение; это называется гетеросинаптическим облегчением. Рассмотрим два примера.
- Первый тип — постсинаптическое облегчение в нейронах симпатических ганглиев. Наряду с другими синаптическими потенциалами там зарегистрированы медленные ВПСП (мВПСП), опосредованные АцХ. Длительность мВПСП может достигать 100 мс. Кроме того, ганглионарная клетка образует синапсы еще и с окончаниями волокон от дофаминергического нейрона. Дофамин, высвобождаемый из этих нервных окончаний, сам по себе не изменяет ионную проводимость постсинаптической мембраны. Однако, действуя в качестве модулятора, он повышает амплитуду мВПСП на несколько часов. Этот эффект обусловлен усилением постсинаптической реакции на АцХ.
P.S. Дофаминовые рецепторы. Различают пять типов рецепторов дофамина, которые через посредство G-белков стимулируют активность аденилатциклазы (рецепторы D1,5), либо блокируют ее (рецепторы D2-4). Дофамин может инициировать симптомы шизофрении, и блокаторы рецепторов дофамина используются при лечении этого заболевания. В нигростриатной системе дофамин осуществляет главную роль в регуляции двигательных функций.
- Второй тип — пресинаптическое облегчение. Оно обнаружено у моллюсков и насекомых, однако, по-видимому, имеет отношение и к физиологии человека. При активации серотонинергических нервных волокон (см. далее) высвобождаемый из них серотонин действует на пресинаптическую мембрану, блокируя К+-каналы. В результате замедляется реполяризация мембраны после потенциала действия.
Более длительная деполяризация нервных окончаний обеспечивает повышенную секрецию медиатора, иными словами, приводит к пресинаптическому облегчению. Благодаря одновременной активации (коактивации) двух синапсов возрастает также эффективность синаптического пути.
- В ЦНС человека серотонин вырабатывается в нейронах ядра шва (nucleus raphe), которые посылают проекции в спинной мозг, мозжечок, таламус, гипоталамус, базальные ганглии, лимбическую систему и кору головного мозга. Для серотонина существуют метаботропные, а также ионотропные рецепторы. Пониженное содержание серотонина или ослабление его действия могут способствовать развитию депрессии. При усилении действия серотонина или стимуляции рецепторов серотонина наблюдается антидепрес-сивный эффект.
1. Суммация. Большинство нейронов несут на себе множество синапсов, потенциалы и токи которых могут суммироваться. Пространственная суммация соответствует объединению реакций, одновременно возникающих в разных синапсах на клетке, временная суммация означает взаимодействие реакций, следующих друг за другом с небольшими интервалами в одном и том же синапсе либо в группе соседствующих синапсов.
От аксонного холмика возбуждение распространяется ретроградно, т. е. к дендритам, где в результате возникают потенциалзависимые Na+- или Са2+-то-ки. Активация дендритов участвует в пластических изменениях синапсов.
2. Торможение. Одна из форм взаимодействия синапсов на отдельном нейроне — это синаптическое торможение. При постсинаптическом торможении тормозные постсинаптические потенциалы (ТПСП) гиперполяризуют мембрану, препятствуя тому, чтобы деполяризация достигла порога возбуждения. В случае пресинаптического торможения аксо-аксонный синапс ингибирует высвобождение медиатора из возбуждающего пресинаптического окончания.
3. Гетеросинаптическое облегчение. При гетеросинаптическом облегчении аксо-аксонный синапс стимулирует высвобождение медиатора из возбуждающего пресинаптического окончания. Другой вариант состоит в том, что один синапс повышает постсинаптическую эффективность другого синапса.
Видео физиология торможения в ЦНС - профессор, д.м.н. П.Е. Умрюхин
- Рекомендуем ознакомиться далее "Механизм высвобождения медиатора в синапсе (нейросекреция) и синаптическое облегчение - с точки зрения физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 9.10.2024