
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Распространение электрических сигналов в мембране нейронов - с точки зрения физиологии человека
Оглавление статьи:- Простой случай: сферическая невозбудимая клетка
- Более сложный случай: цилиндрическая невозбудимая структура (кабель)
- Непрерывное проведение импульсов в немиелинизированных нервных волокнах
- Миелинизация нервных волокон обеспечивает высокую скорость проведения
- Рассеянный склероз
- Резюме
- Список использованной литературы
У многих нейронов удалены друг от друга входные синапсы, которые опосредуют их активацию, и выходные синапсы, направляющие сигналы к клеткам-мишеням. Максимальная длина дендрита может доходить до нескольких миллиметров, максимальная длина аксона — до 2 м. Отсюда возникает вопрос: как электрические сигналы нервных клеток способны преодолевать такие большие расстояния?
а) Простой случай: сферическая невозбудимая клетка. Дендриты и аксоны можно представить как «биологический кабель» из большого числа параллельно соединенных электрических элементов (емкостных и резистивных).
Наиболее простой пример распространения электрического сигнала — это его распространение по сферической невозбудимой клетке (рис. 1). Инъекция прямоугольного импульса постоянного тока сопровождается экспоненциальным нарастанием электрического заряда клеточной мембраны, а выключение тока, наоборот, экспоненциальным уменьшением заряда.
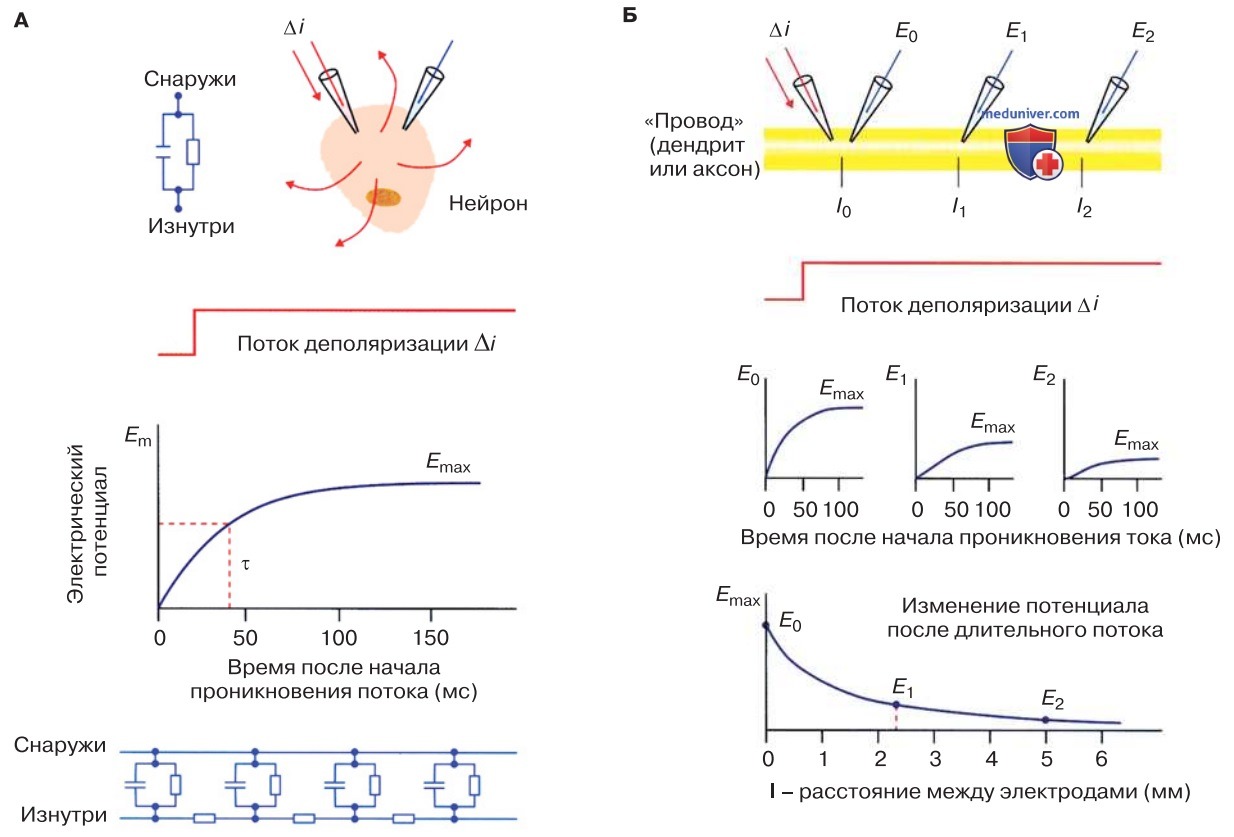
1. Количественный анализ. Если представить себе, что в некоторый момент времени заряд одинаков во всех участках мембраны (мембрана изо-потенциальна), то несложно провести количественный анализ изменений электрического потенциала.
Клеточная мембрана состоит из конденсатора (емкости) и резистора (сопротивления) (рис. 1), причем емкость соответствует липидному бислою, а сопротивление — проницаемым трансмембранным каналам (например, двупоровым, иными словами, 2Р-доменным К+-каналам).
Мембранный потенциал Е изменяется экспоненциально как функция времени t, т. е. E(t) = Emax(1 - e-t/τ) (емкость заряжается) либо E(t) = Emaxe-t/τ (емкость разряжается); Emax — максимальное изменение напряжения, τ -постоянная времени мембраны.
Постоянная времени мембраны — это время нарастания мембранного потенциала до уровня 1-1/е = 63% (е = число Эйлера) от своего конечного (максимального) значения (в фазу нарастания заряда) или его падения до уровня 1/е = 37% (в фазу уменьшения заряда; рис. 1, внизу).
- В простом случае (в сферической клетке) постоянная времени мембраны представлена как τ = Rm * Cm, где Rm — сопротивление мембраны, Cm — емкость мембраны. Часто обе переменные нормализованы на единицу площади поверхности мембраны. Тогда речь идет о специфическом сопротивлении мембраны (Ом * см2) и специфической емкости мембраны (мкФ/см2).
б) Более сложный случай: цилиндрическая невозбудимая структура (кабель). Проведение потенциалов действия по немиелинизированным аксонам — это непрерывный процесс.
Более сложен случай, когда электрический ток входит через мембрану цилиндрического отростка — дендрита или аксона («кабельная модель»). Здесь при таких же свойствах мембраны, что и в сферической клетке, временной ход изменения мембранного потенциала отличается: он существенно зависит от места пропускания тока и не описывается экспонентой. По сравнению с электрическими характеристиками сферической клетки (см. выше) наблюдаются следующие особенности.
- Непосредственно в месте инъекции тока (источник тока) мембранный потенциал нарастает быстрее, чем в сферической клетке.
- По мере удаления от источника тока электрический потенциал нарастает медленнее (кривая становится более плоской) и начинается с некоторым отставанием от момента инъекции тока.
- По мере удаления от источника тока максимальная амплитуда тока постепенно снижается. В отличие от процессов в электрическом кабеле (например, в изолированном медном проводе) в биологическом кабеле проведение сигнала происходит с некоторым отставанием по времени, а также с потерей амплитуды.
1. Количественный анализ. Количественный анализ изменений мембранного потенциала кабельной структуры существенно сложнее. Кабель рассматривается как цепь конденсаторов и резисторов — элементов, моделирующих двойной липидный слой и проницаемые каналы соответственно, которые соединены друг с другом через резисторы (моделирующие цитоплазму) (рис. 1).
В такой пассивной системе между максимальной амплитудой сдвига потенциала (Emax) и местом пропускания тока (х) существует следующая взаимосвязь: Emax(x) = = Emax(0)e-x/τ. Постоянная длины мембраны (Д) — это свойство кабеля; она соответствует расстоянию, на котором Emax снижается до 37% (на 1/е) от амплитуды потенциала в месте инъекции тока.
Величина постоянной длины мембраны позволяет судить о радиусе распространения электрического сигнала в биологическом кабеле (рис. 1, внизу); процесс электротонический.
- Постоянная (константа) длины рассчитывается по формуле:
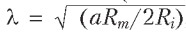 ,
,где а — радиус кабеля, Ri — внутриклеточное сопротивление. Точнее, Ri — это удельное внутриклеточное сопротивление, формула которого учитывает длину и площадь поперечного сечения кабеля (Ом * см, или Ом * мм2/м). Отсюда два важных вывода.
При постоянном значении Rm и Ri; константа длины λ пропорциональна корню квадратному из радиуса кабеля (√а); т. е., чем больше радиус, тем больше постоянная длины кабеля, и, следовательно, зона действия электрического сигнала.
При постоянном значении радиуса (а) константа длины λ пропорциональна корню квадратному из отношения Rm/Ri. Таким образом, чем больше сопротивление мембраны по отношению к внутриклеточному сопротивлению, тем (опять же) больше зона действия электрического сигнала. Две основные характеристики биологического кабеля — постоянная времени мембраны и постоянная длины — определяют скорость пассивного проведения, которая рассчитывается как ~ 2λ/τ.
Поскольку постоянная длины Д пропорциональна корню квадратному из радиуса кабеля (√а), скорость пассивного проведения тоже должна быть пропорциональна √а. Чем больше радиус биологического кабеля, тем выше скорость пассивного проведения. Для кабельных характеристик дендритов приняты следующие приблизительные значения: постоянная времени мембраны τ ~ 10 мс, постоянная длины мембраны λ ~ 1 мм, скорость пассивного проведения 2λ/τ ~ 0,2 м/с.
Кабельные свойства дендритов играют важную роль в процессе интеграции сигналов. Во многих случаях постоянная времени мембраны может обеспечить уменьшение длительности возбуждающих постсинаптических потенциалов (ВПСП), а также укорочение временного окна для суммации синаптических потенциалов. От константы длины мембраны дендрита зависит степень демпфирования ВПСП при его распространении по дендриту к соме нейрона.
Поскольку ВПСП имеет транзиторный характер, эффект его ослабления оказывает более существенное влияние, чем в случае сдвига потенциала, вызванного прямоугольным импульсом тока (см. выше); коэффициент удовлетворяет константе длины Д. Следовательно, синаптический сигнал, возникший в дистальной части дендрита, имеет фактор ослабления < 1/е ~ 37%.
В отличие от мембраны большинства дендритов мембрана аксонов (а также некоторых дендритов) ведет себя как активная структура, поскольку обладает NaV- и KV-каналами. По отношению к аксонам кабельная теория применима только при слабых стимулах; при более интенсивных стимулах, вызывающих ПД, распространение возбуждения определяется свойствами и распределением потенциалуправляемых ионных каналов, а не только кабельными характеристиками волокна.
в) Непрерывное проведение импульсов в немиелинизированных нервных волокнах. В миелинизированных аксонах проведение ПД осуществляется быстро благодаря тому, что возбуждение «перескакивает» от одного перехвата к следующему практически без декремента («сальтаторное проведение»).
Инициация ПД происходит в структурах нервной клетки, имеющих особенно высокую плотность NaV-каналов — в аксонном холмике или начальном сегменте аксона. Если аксон немиелинизированный, то ПД распространяется непрерывно (рис. 2). При этом между возбужденным и соседним невозбужденным участком мембраны возникает ток, который деполяризует невозбужденную область мембраны и, следовательно, активирует находящиеся там NaV-каналы. Развивается сильная деполяризация, которая приводит к распространению возбуждения по всему аксону (рис. 2).
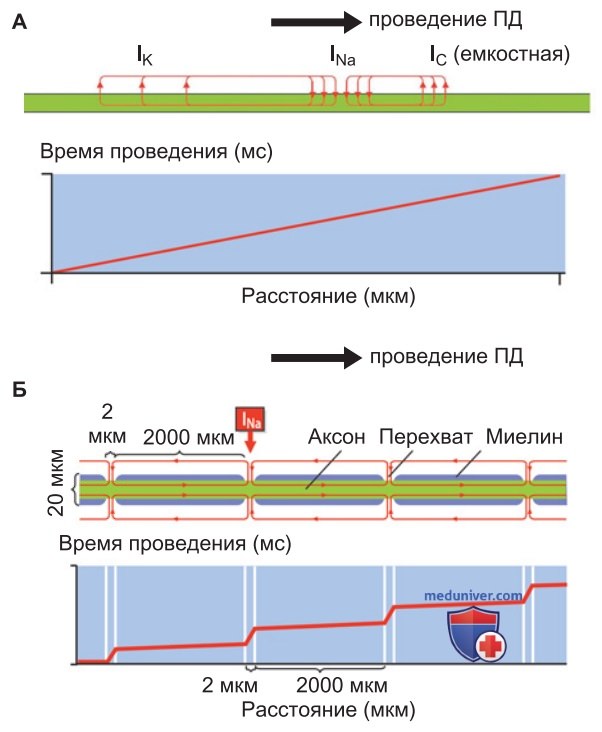
При непрерывном проведении ПД общий профиль трансмембранного тока складывается из трех компонентов:
- зоны входящего тока в центральной части ПД (NaV-каналы);
- зоны выходящего тока перед ПД (разрядка емкости мембраны);
- зоны выходящего тока позади ПД (KV-каналы).
Между зонами входящего и выходящего токов образуются локальные петли тока, которые проходят через аксоплазму и внеклеточную жидкость.
Скорость активного непрерывного проведения. Так же как в случае пассивного проведения, при активном непрерывном проведении скорость пропорциональна корню квадратному из радиуса волокна √а. Главная закономерность такова: толстое волокно — быстрое проведение, тонкое волокно -медленное проведение. Особенно толстое нервное волокно — так называемый «гигантский аксон» головоногих моллюсков, его диаметр ~1 мм.
В подтверждение отмеченной выше закономерности, он обладает достаточно высокой скоростью проведения (~20 м/с). В организме человека диаметр немиелинизированных аксонов равен ~1 мкм. При этом скорость проведения не превышает ~1 м/с (табл. 2).
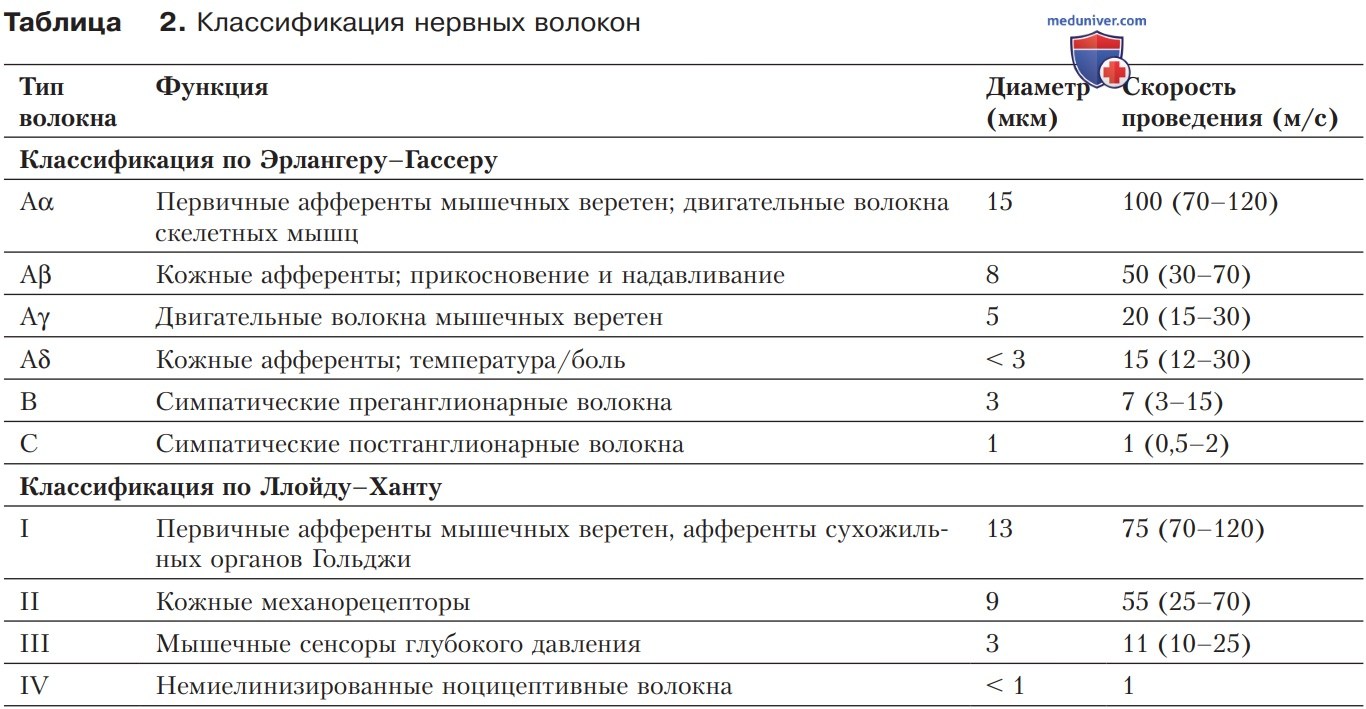
г) Миелинизация нервных волокон обеспечивает высокую скорость проведения. Высокая скорость сальтаторного проведения объясняется высокой плотностью распределения NaV-каналов в мембране перехватов Ранвье, а также особенностями проводимости межперехватных участков (время проведения в которых практически равно нулю).
В миелинизированных нервных волокнах возбуждение распространяется гораздо быстрее, чем в немиелинизированных (рис. 2). Такая высокая скорость проведения возможна благодаря сочетанию активной и пассивной формы проведения. В отличие от немиелинизированного аксона, где активные процессы (активация NaV-каналов) и пассивные (зарядка емкости мембраны) происходят одновременно, в миелинизированных волокнах они отделены друг от друга как во времени, так и в пространстве. Активные процессы ограничиваются перехватами Ранвье.
Пассивные механизмы, напротив, сосредоточены в межперехватных сегментах, формирующихся в процессе миелинизации аксона.
1. Различия между перехватами Ранвье и межперехватными сегментами. Электрические свойства межперехватного участка и перехвата принципиально различаются. Это обусловлено особенностями распределения потенциалуправляемых ионных каналов. NaV-каналы концентрируются в мембране перехватов (более тысячи на 1 мкм-2), но практически отсутствуют в околоперехватных и межперехватных сегментах. Там сосредоточены KV-каналы, которые почти лишены перехватов.
Вследствие недостаточного числа KV-каналов в перехвате Ранвье ПД возникают только во время инактивации NaV-каналов и потенциалнеуправляемых (каналов утечки) двупоровых К+-каналов с 2-Р доменами.
- Пассивные электрические свойства межперехватного сегмента и перехвата Ранвье существенно различаются.
- В межперехватном участке специфическая емкость примерно в 250 раз меньше, чем в перехвате Ранвье, поскольку внутриклеточное пространство отделено от внеклеточного миелиновой оболочкой. Хотя межперехватные сегменты намного длиннее, чем перехваты, абсолютные значения их емкости близки (Смежперехват = 2-4 пФ, Сперехват = = 0,6-1 пФ).
- В межперехватном участке специфическое радиальное сопротивление в ~ 8000 раз выше, чем в перехвате. Свойства обоих участков благоприятно влияют на проведение ПД через межперехватный сегмент. В самом деле, поскольку в межперехватном сегменте потеря заряда невелика, к следующему перехвату поступает большой заряд.
2. Сальтаторное проведение возбуждения. Средняя скорость проведения возбуждения в миелинизированных волокнах может составлять в зависимости от типа волокна до 100 м/с. Такая высокая скорость проведения — важное следствие миелинизации. Когда ПД достигает перехвата Ранвье, возникает ток, который распространяется к следующему перехвату, вызывая деполяризацию. Ввиду особых свойств межперехватного сегмента (высокое сопротивление и низкая емкость) деполяризация соседних перехватов осуществляется эффективно и быстро (возбуждение «перескакивает» от перехвата к перехвату).
Поэтому проведение возбуждения в миелинизированных аксонах получило название сальтаторного (скачкообразного). Общие расчеты показывают, что ~50% времени проведения возбуждения в перехвате Ранвье приходится на активные процессы (повышение положительного заряда мембраны, активация NaV-каналов), тогда как еще 50% относятся к пассивному проведению через межперехватные сегменты (рис. 2).
3. Классификация нервных волокон. Нервные волокна классифицируются по наличию миелинизации, диаметру и скорости проведения возбуждения, которая зависит от первых двух факторов. Существуют классификации Эрлангера-Гассера и Ллойда-Ханта (для сенсорных нервов) (табл. 2).
Быстрое сальтаторное проведение возбуждения в миелинизированных волокнах эффективно с точки зрения энергозатрат. Согласно приблизительным расчетам, при одной и той же скорости проведения при каждом ПД для толстого немиелинизированного волокна по сравнению с миелинизированным нужна гораздо более значительная площадь поперечного сечения и в несколько тысяч раз больше метаболической энергии.
Как при непрерывном, так и при сальтаторном проведении импульс может распространяться по аксону в обоих направлениях. Какое направление считать физиологическим, зависит исключительно от места генерации ПД. Проведение в физиологическом направлении называется ортодромным, проведение в обратном направлении — антидромным. Антидромное проведение возможно, например, при электрической стимуляции.
1) Симптомы и признаки. Рассеянный склероз — прогрессирующее аутоиммунное заболевание нервной системы, которое может проявляться как расстройство чувствительности (покалывание и онемение рук и ног, болевые синдромы), зрительные нарушения, спастические параличи, усиление тонических мышечных рефлексов на растяжение, нарушения координации движений (атаксия).
2) Патогенез. Происходит демиелинизация аксонов в головном и спинном мозге. Из-за патологических изменений структуры миелиновой оболочки нарушается сальтаторное проведение возбуждения, так что снижаются скорость и эффективность передачи сигналов. Поскольку демиелинизация может происходить в различных отделах головного мозга, симптоматика заболевания разнообразна.
д) Резюме. Отросток нервной клетки можно рассматривать как «биологический кабель» — электрическая цепь из резистивных и емкостных элементов. Определяющая роль в интеграции и проведении синаптических потенциалов от дендритов к соме принадлежит постоянной времени и постоянной длины мембраны. Проведение ПД по аксонам представляет собой активный процесс (активация NaV-каналов).
От точки возникновения ПД распространяется с высокой скоростью вдоль всего аксона нервной клетки к синаптическим окончаниям. В немиелини-зированных волокнах проведение осуществляется непрерывно (со скоростью ~1 м/с), а в миелинизированных волокнах это сальтаторный процесс, скорость которого может достигать 100 м/с.
Высокая скорость сальтаторного проведения обусловлена сосредоточением NaV-каналов в перехватах Ранвье, а также особыми электрическими свойствами межперехватных сегментов.
Видео физиология синапса и нерва - профессор, д.м.н. П.Е. Умрюхин
- Рекомендуем ознакомиться далее "Ритмическая активность и кодирование информации в нервной системе - с точки зрения физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 10.9.2024