
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Строение потенциалуправляемых катионных каналов клетки - с точки зрения физиологии человека
Оглавление статьи:- Топология и структурная организация
- Классификация катионных каналов
- Резюме
- Список использованной литературы
а) Топология и структурная организация. Ионные каналы состоят из порообразующих α-субъединиц и дополнительных β-субъединиц; структура белка-каналоформера определена аминокислотной последовательностью и мембранной топологией этих компонентов.
1. Мембранная топология. Аминокислотная последовательность (первичная структура) белка-каналоформера содержит гидрофобные и гидрофильные участки (рис. 1). От них зависит размещение белка в клеточной мембране (мембранная топология) и в итоге — характер сворачивания белковой молекулы, т. е. его третичная структура.
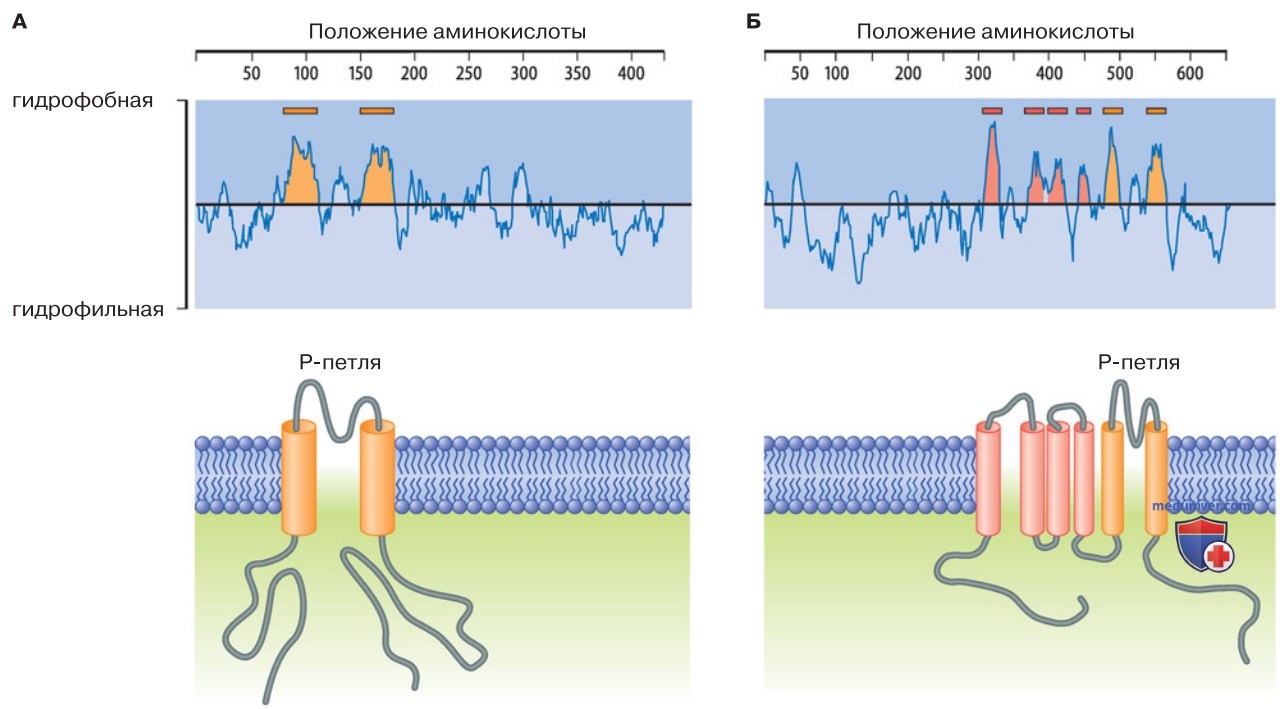
- Гидрофобные участки (в основном вторичные структуры) обычно имеют конфигурацию α-спиралей, пронизывающих липидный бислой клеточной мембраны.
- Гидрофильные участки, в том числе N-и C-концевые участки полипептидов, находятся в водной среде внутриклеточного и внеклеточного пространств либо поры, проводящей ионы (рис. 1).
Количество гидрофобных и гидрофильных участков может быть разным, причем эволюционно более древними являются, видимо, формы с двумя или шестью трансмембранными сегментами (двух- или шестисегментные каналы).
2. Структура поры ионного канала. Порообразующий белок двух- или шестисегментного канала — это единая молекула из нескольких полипептидов (четвертичная структура). Методом рентгеноструктурного анализа кристаллизованного белка была изучена атомная структура относительно простого двухсегментного калиевого канала (рис. 2).
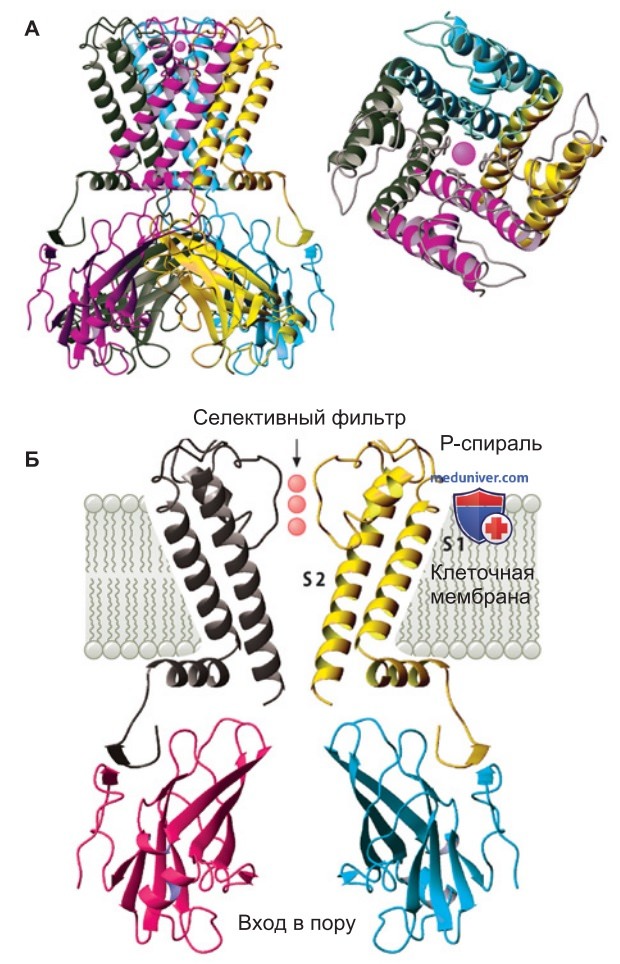
Этот калиевый канал представляет собой тетрамер; четыре его субъединицы располагаются вдоль оси симметрии белка, окружая пору. Со стороны цитоплазмы стенка поры образована С-концом трансмембранной спирали (внутренней спирали S2), а с внеклеточной стороны — соединительным доменом (Р-петлей, или Р-доменом) двух трансмембранных сегментов (рис. 1). Молекула канала отделена от липидного матрикса N-концом трансмембранной спирали (наружной спирали S1), которая располагается за внутренней спиралью и слегка наклонена по отношению к ней.
С цитоплазматической стороны устье поры образуют N- и C-концы четырех субъединиц канала (рис. 2).
3. Селективный фильтр. Селективный фильтр K+-канала — самый узкий участок поры, который находится недалеко от наружного устья; он образован С-концом Р-петли и стабилизирован короткой спиралью поры (рис. 3).
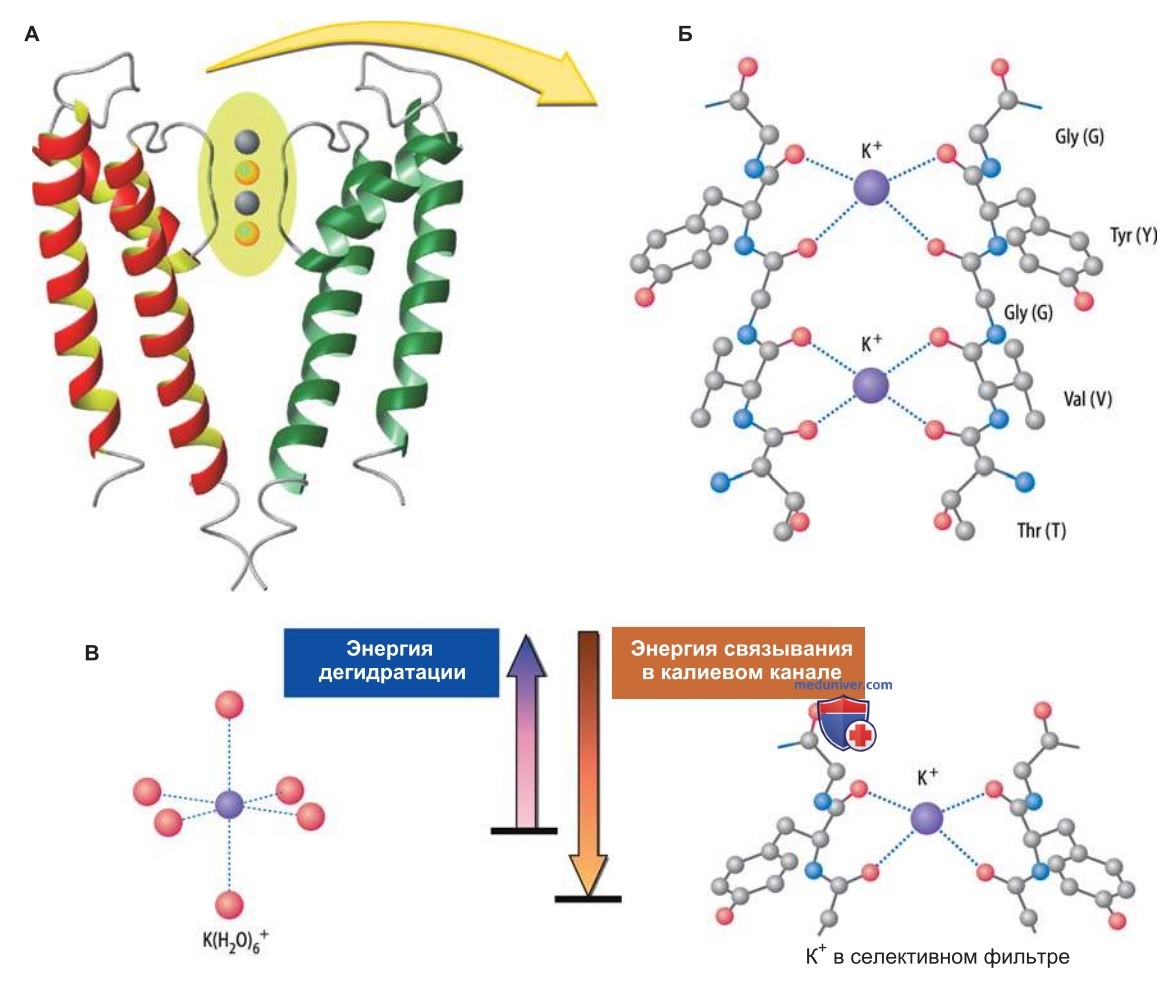
В K+-канале любого типа стенка селективного фильтра содержит характерную последовательность аминокислот (глицин—тирозин—глицин; GTG-мотив); ее особенность в том, что карбоксильные группы остатков тирозина и внутренних остатков глицина четырех субъединиц каналоформера образуют кольцевую структуру (рис. ниже), идеально заменяющую гидратную оболочку для K+, но не для Na+ или Li+.
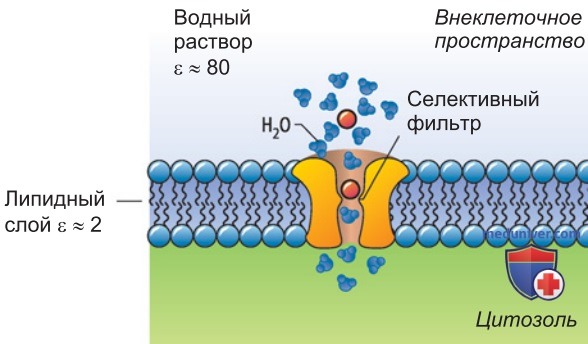
Такой «заместитель гидратной оболочки» обеспечивает высокую селективность K+-канала, т.е. его способность пропускать более крупные ионы K+ (радиус 1,33 А), но не мелкие ионы Na' (радиусом 0,95 А) или Li+ (радиусом 0,6 А) (рис. 3).
- Структурные основы проницаемости и селективности калиевого канала. Рентгеноструктурное исследование кристаллов двухсегментного К+-канала показало, что вследствие сниженного барьера Борна в этом канале возможен более значительный поток ионов. Прежде всего здесь очень короткий селективный фильтр, так что внутриклеточная среда практически соприкасается с внеклеточной и энергия, необходимая для проникновения иона, уменьшается до минимума.
Кроме того, при электростатическом взаимодействии между атомом кислорода карбоксильной группы и ионом высвобождается дополнительная энергия для перехода иона через мембрану (рис. 3). Другую важную особенность структуры канала составляет дипольный момент спиралей поры, обусловленный неравным распределением заряда α-спиралей: благодаря особому расположению вокруг поры четырех спиралей возникает отрицательный потенциал в селективном фильтре, способствующий входу катионов и выталкиванию анионов.
б) Классификация катионных каналов. В зависимости от аминокислотной последовательности и мембранной топологии катионные каналы подразделяются на классы, семейства и подсемейства.
1. Классы каналов. Порообразующая субъединица большинства ионных каналов либо состоит из двух последовательных трансмембранных сегментов, соединенных Р-доменами, либо представляет собой аналогичную структуру. Гены известных каналов принято подразделять исходя из количества подобных структур, а также числа и свойств прочих трансмембранных сегментов (рис. 4). Наиболее простое эволюционное расширение структуры двухсегментного канала — это шестисегментный канал. В пределах этого класса каналов выделено более 90 (!) различных генов.
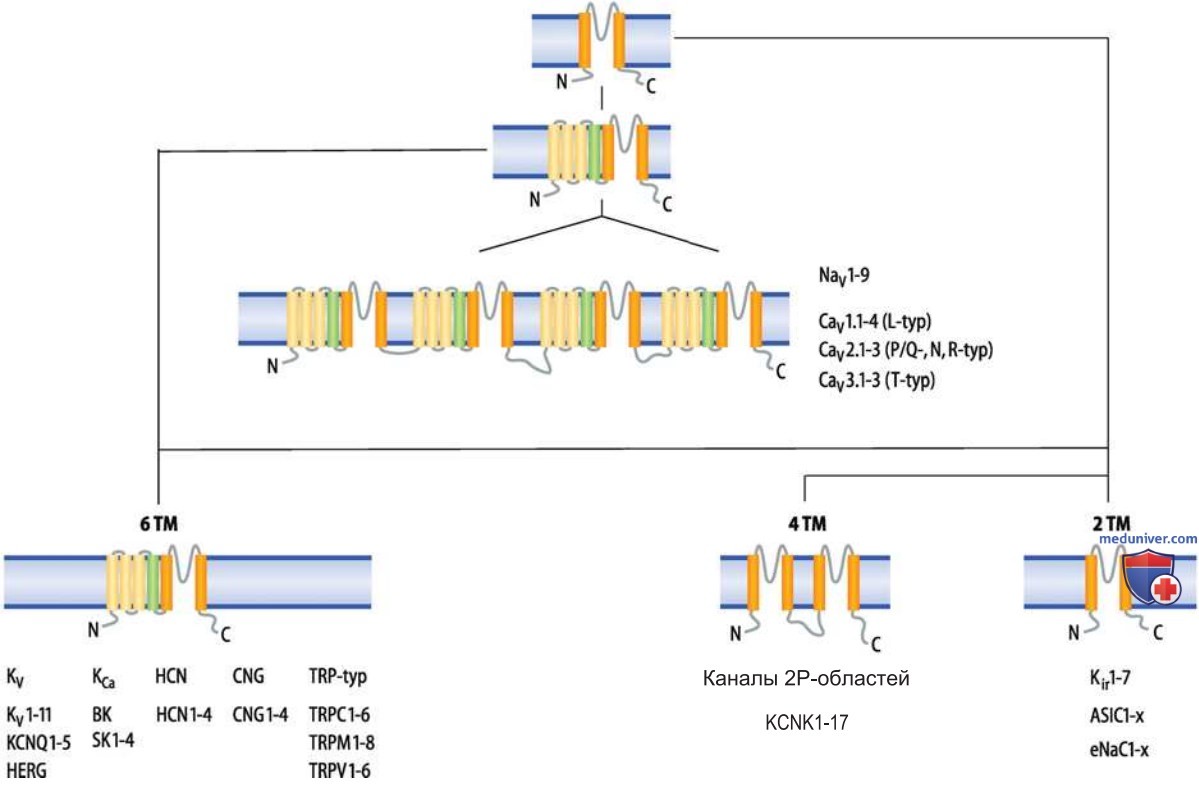
Что касается четырех дополнительных трансмембранных сегментов, то обнаружена следующая особенность первичной структуры самой последней спирали S4: каждая третья позиция внутри этой спирали занята положительно заряженной аминокислотой — аргинином или лизином, так что сегмент S4 при физиологическом уровне рН получает положительный заряд. Сегмент S4 служит в каналах «сенсором» — детектором изменений мембранного потенциала; этот механизм присутствует во всех потенциалуправляемых ионных каналах.
С точки зрения модульного принципа каналы других классов можно рассматривать как объединения двух- и шестисегментных субъединиц (рис. 4). Следовательно, калиевые каналы с двумя Р-доменами являются системой из двух двухсегментных субъединиц, а потенциалуправляемые натриевые и кальциевые каналы (NaV- и CaV-каналы) — объединения из четырех шестисегментных субъединиц.
Подобная система из четырех объединенных шестисегментных субъединиц, кодируемая одним геном, соответствует альтернативному принципу формирования структуры NaV- и CaV-каналов; в то время как двух-, четырех- и шестисегментные каналы построены из четырех субъединиц по «принципу 4 х 1», молекулы NaV- и CaV-каналов образованы одной субъединицей по «принципу 1 х 4».
2. Семейства и подсемейства каналов. На основе сходства аминокислотных последовательностей каналы можно разделить на семейства и подсемейства (рис. 4). Примеры семейств — потенциал-управляемые калиевые каналы (KV), кальций-управляемые калиевые каналы (КCa), калиевые каналы внутреннего выпрямления (Kir); примеры подсемейств — KV1-, SK-, а также Кir-2-каналы.
Подобное подразделение играет важную роль в развитии представлений об архитектуре каналов, поскольку двух- и шестисегментные каналы необязательно должны включать в себя четыре идентичные субъединицы (такие объединения называются гомомерами), но могут также быть гетеромерами, т. е. состоять из различных молекул. Однако гетеромультимерность возможна только в пределах а-субъединиц одного подсемейства, но не среди представителей различных семейств или классов ионных каналов.
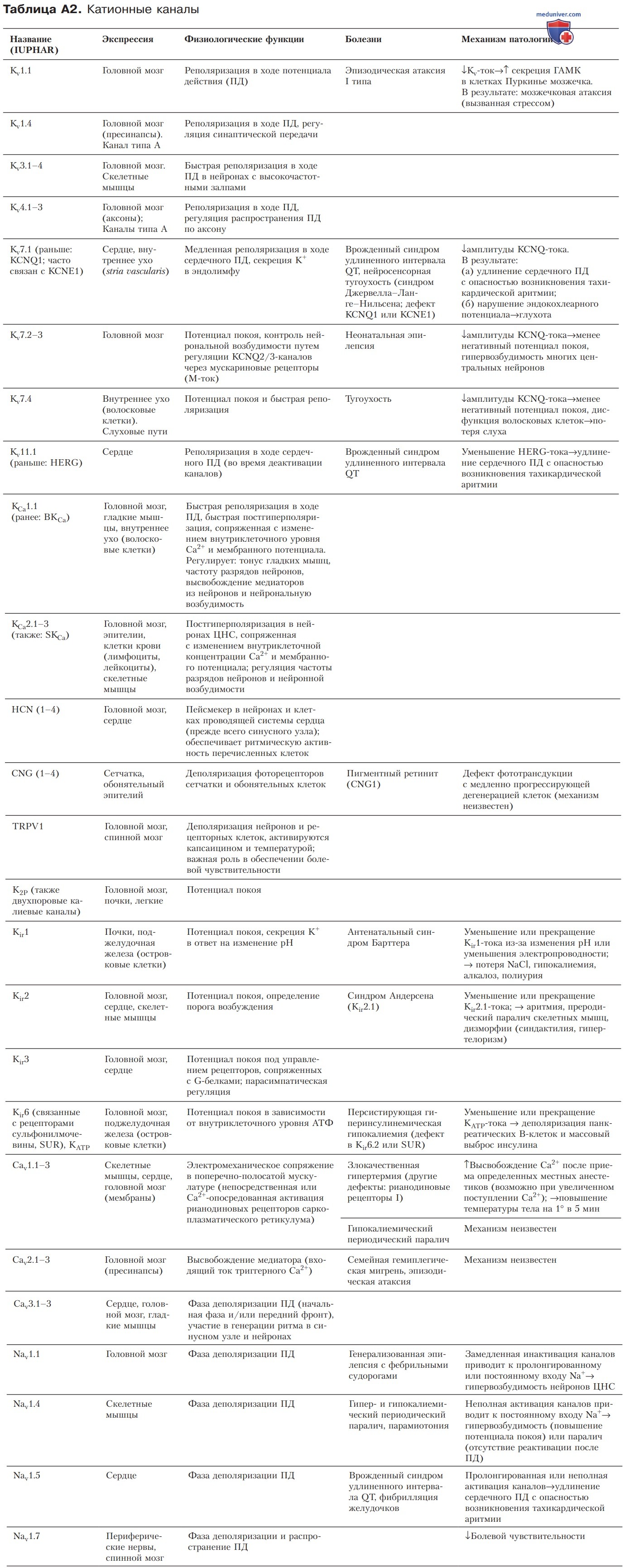
Представленная классификация генов, кодирующих каналы различного типа, учитывает прежде всего структуру и основные функциональные характеристики каналов. Физиологические свойства обобщены в табл. А2 и более подробно описаны в отдельных статьях на сайте (просим Вас пользоваться формой поиска по сайту выше), посвященных тканям и органам, где функционирует соответствующий каналообразующий белок.
3. Дополнительные субъединицы. Наряду с перечисленными порообразующими субъединицами многие каналы содержат ассоциированные белки, которые не входят в состав самой поры и поэтому обозначаются как дополнительные элементы.
В структурном отношении дополнительными элементами могут быть интегральные мембранные белки: с гидрофобными трансмембранными сегментами (например, β-субъединицы NаV-каналов или калиевых каналов KCNQ- или ВК-типа (см. табл. А2)) либо преимущественно гидрофильные цитоплазматические белки (например, β-субъединицы Куканалов или CaV-каналов). Они соединены с порообразующей α-субъединицей посредством дисульфидных связей и гидрофобных взаимодействий.
Функциональная роль дополнительных субъединиц разнообразна: одни из них управляют переключением состояния канала (так называемый воротный механизм, см. далее), другие субъединицы модулируют проводимость канала, участвуют в процессинге белков, обеспечивают локализацию или стабилизируют ориентацию α-субъединиц в мембране.
1. Строение потенциалуправляемых катионных каналов. Катионные каналы состоят из порообразующих α-субъединиц и акцессорных (дополнительных) субъединиц (имеется в виду четвертичная структура). Каждая α-субъединица в катионном канале включает в себя гидрофобные и гидрофильные участки, от которых зависит характер сворачивания трансмембранного белка (мембранная топология, третичная структура); число годрофобных и гидрофильных участков и их расположение определяются аминокислотной последовательностью белка (первичной структурой).
Гидрофобные сегменты в основном соответствуют пронизывающим мембрану а-спиралям (вторичной структуре). Гидрофильные сегменты находятся в водной среде внеклеточного и внутриклеточного пространств.
Функциональный белок канала формируется в результате объединения четырех α-субъединиц (тетрамерная структура), лишь белки NaV-каналов и CaV-каналов образованы одной α-субъединицей из четырех мембранных доменов (псевдотетрамерная структура).
Пора канала расположена вдоль оси симметрии белка. Самое узкое место поры, так называемый селективный фильтр, находится вблизи внеклеточного устья.
2. Классификация. Известно, что геном человека содержит большое число генов, кодирующих α-субъединицы потенциалуправляемых катионных каналов. На основании сходства их первичной аминокислотной последовательности и третичной структуры, т. е. топологии мембраны (числа гидрофобных сегментов), белки катионных каналов принято подразделять на различные классы, семейства и подсемейства.
- Рекомендуем ознакомиться далее "Воротные механизмы катионных каналов клетки - с точки зрения физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 10.9.2024