
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Лигандактивируемые ионные каналы клетки - с точки зрения физиологии человека
Оглавление статьи:- Строение ионных каналов возбуждающих рецепторов
- Функциональные свойства ионных каналов возбуждающих рецепторов
- Структурные и функциональные свойства ионных каналов тормозных рецепторов
- Резюме
- Список использованной литературы
а) Строение ионных каналов возбуждающих рецепторов. Лигандактивируемые ионные каналы возбуждающих рецепторов (ионотропных рецепторов) состоят из четырех или пяти субъединиц.
Важнейшим способом активации каналов, наряду с изменением мембранного потенциала, является связывание внеклеточного медиатора (лиганда) с каналом. Ионные каналы, активируемые таким образом, представляют собой лигандактивируемые каналы, или ионотропные рецепторы. Наименование конкретного типа канала образуется от названия активирующих лигандов (агонистов); следовательно, канал, управляемый ацетилхолином (АцХ), известен как ионотропный ацетилхолиновый рецептор.
В отличие от потенциалуправляемых каналов лигандактивируемые каналы находятся в основном в постсинаптических структурах, поскольку медиаторы способны встретиться с ними только там.
Существует множество генов, кодирующих ионотропные рецепторы. На основании аминокислотных последовательностей и структуры белков ионотропные рецепторы можно разделить на классы, семейства и подсемейства. Дальнейшее подразделение в большей мере учитывает физиологические функции каналов, иными словами, тип ионов, проникающих через канал. Так, лигандактивируемые катионные каналы определяются как каналы возбуждающих рецепторов, а анионные каналы — как каналы тормозных рецепторов.
1. Ионные каналы возбуждающих рецепторов. Важнейшими возбуждающими медиаторами организма млекопитающих считаются глутамат и АцХ; отсюда особое внимание к таким возбуждающим рецепторам как ионотропные глутаматные рецепторы (iGluR) и ионотропные АцХ-рецепторы (получившие также название никотиновые ацетилхолиновые рецепторы, nAchR, благодаря их способности активироваться никотином). В соответствии с их селективными агонистами iGluR подразделяются на NMDА-рецепторы (N-метил-D-аспартатные), АМРА-рецепторы (α-амино-3-гидрокси-5-метил-4-изоксазолпропионатные) и каинатные рецепторы.
Физиологическая роль nAchR особенно значима в периферической нервной системе и скелетной мускулатуре (в нервно-мышечных синапсах, или концевых пластинках), а iGluR — в центральной нервной системе (ЦНС).
2. Строение ионных каналов возбуждающих рецепторов. Что касается мембранной топологии, то каналообразующий белок рецепторов двух типов (iGluR и nAchR) состоит из четырех гидрофобных сегментов, которые, однако, образуют каналы, несколько различающиеся по структуре (рис. ниже). В случае iGluR три сегмента (М1, М3 и М4) — это трансмембранные домены, а сегмент М2 представляет собой Р-петлю на уровне мембраны и участвует в структуре поры аналогично Р-доменам калиевых и натриевых каналов. Длинный N-концевой домен белка iGluR находится во внеклеточном пространстве, а короткий С-конец — с цитоплазматической стороны мембраны.
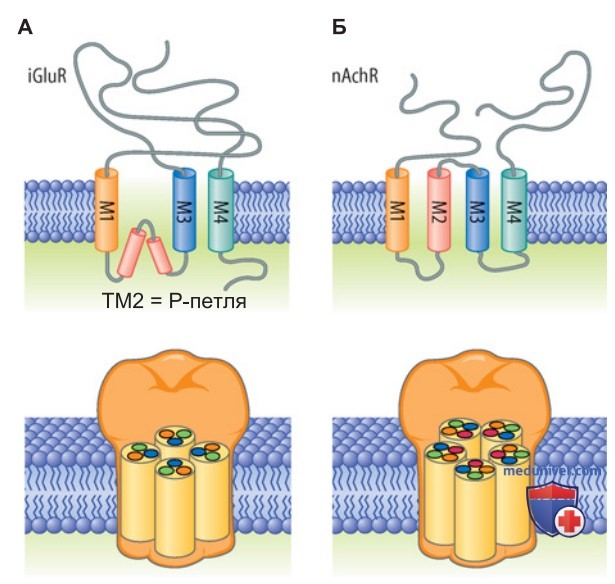
В белке nAchR все четыре гидрофобных сегмента, напротив, являются трансмембранными доменами, так что N- и С-концевые домены расположены во внеклеточном пространстве.
Таким особенностям четвертичной структуры того и другого рецептора соответствует стехиометрия их структурных компонентов, а также строение участка связывания с лигандом. Глутаматные рецепторы (iGluR) — это тетрамеры (рис. выше), которые в зависимости от их типа состоят из четырех идентичных или четырех различных субъединиц. Так, iGluR NMDA-типа — гетеротетрамеры с субъединицами NR1 и NR2, iGluR АМРА-типа — гомо- или гетеротетрамеры с субъединицами GluR1-4, а каинатные рецепторы — гомо- или гетеротетрамеры с субъединицами GluR5-7 и KA1-2 (табл. 1).

Все субъединицы iGluR обладают участками связывания глутамата, которые образованы N-концевым доменом и соединительным элементом трансмембранных сегментов М3 и М4.
Напротив, nAch-рецепторы обычно состоят из пяти различных субъединиц (являются пентамерами) (рис. выше). При этом nAchR скелетной мышцы представляют собой гетеропентамеры, состоящие из двух α1-субъединиц, а также одной β-, γ- (либо ε-) и δ-субъединицы; nAchR нервной системы — пентамеры из двух или трех а-субъединиц (α2-10) и трех или двух β-субъединиц (β2-4).
Согласно современным данным, nAchR каждого типа имеет два участка связывания с агонистами, которые чаще всего принадлежат а-субъединице. Пора канала nAchR образована М2-сегментами пяти субъединиц, а также смежными участками белка (рис. выше).
б) Функциональные свойства ионных каналов возбуждающих рецепторов. Ионотропные рецепторы активируются при связывании с внеклеточными лигандами/ медиаторами; возбуждающие глутаматные и ацетилхолиновые рецепторы представляют собой неселективные катионные каналы.
1. Воротные механизмы. Несмотря на отмеченные выше различия структуры рецепторных белков, молекулы iGluR- и nAchR-рецепторов близки по своим функциональным свойствам, механизмам переключения состояний и пропускания ионов. Так же как потенциалзависимые каналы при гиперполяризации мембраны, каналы рецепторов в отсутствие агониста находятся в закрытом состоянии (С-состоянии). При этом они могут активироваться в результате связывания с агонистом — глутаматом (для рецепторов NMDA-типа дополнительно требуется связывание с глицином) либо с ацетилхолином.
Взаимодействие агониста с рецептором обеспечивает, аналогично эффекту смещения спирали S4, поступление энергии к белку канала: при связывании с агонистом изменяется конформация участка связывания и его окружения; реорганизуется структура порообразующих сегментов белка; канал открывается (переходит в О-состояние). АМРА-рецепторы и nAchR-рецепторы скелетной мышцы, а также в некоторых нейронных nAchR-рецепторах открываются менее чем на одну миллисекунду, тогда как в других рецепторах, например в NMDA-рецепторе, открытое состояние длится 10 и более миллисекунд.
Открытый канал может снова закрыться, причем для этого существует два способа: деактивация, когда агонист диссоциирует от участка связывания, или же десенситизация посредством инактивации (перехода канала в I-состояние), когда сохраняется связь лиганда с рецептором. Деактивация заканчивается в течение миллисекунд, тогда как продолжительность инактивации может быть очень разной — от нескольких миллисекунд (в nAch-рецепторах скелетной мышцы или в АМРА-рецепторах) до нескольких сотен миллисекунд.
2. Проницаемость каналов. Выше уже отмечалось сходство iGlu- и nAch-рецепторов в отношении проницаемости их ионных каналов. В принципе оба канала пропускают мелкие одновалентные катионы, прежде всего Na+ и K+. В физиологических условиях входящий поток Na+ больше, чем одновременный выходящий поток K+, что обусловлено более значительной движущей силой (см. выше) и относительно более выраженной селективностью Na+-каналов. В результате активация рецепторов как iGlu-, так и nAch-типа приводит к деполяризации постсинаптической мембраны, т. е. к возбуждению постсинаптической клетки.
Некоторые nAch-рецепторы, а также iGlu-рецепторы NMDA-типа способны пропускать не только мелкие одновалентные ионы, но и двухвалентный Са2+, в то время как двухвалентный Mg2+ задерживается и остается «подвешенным» к селективному фильтру, тем самым блокируя пору канала (см. далее).
Наряду с iGluR и nAchR существуют и другие возбуждающие рецепторы, однако их функциональная роль менее значима. К ним относятся:
- ионотропные моноаминовые рецепторы (рецепторы серотонина, или 5-гидрокситриптамина -5-НТ3-рецепторы), которые по структуре близки к nAchR;
- ионотропные АТФ-рецепторы (Р2Х-рецепторы);
- рецепторы с протонными (Н+-) каналами (ASIC), сходные по структуре с двухсегментными белками-каналоформерами двух рассмотренных выше рецепторов.
в) Структурные и функциональные свойства ионных каналов тормозных рецепторов. Лигандактивируемые тормозные ионотропные рецепторы представляют собой анионные каналы-пентамеры; их активируют медиаторы ГАМК и глицин.
1. Строение. Важнейшие тормозные медиаторы ЦНС — это аминокислоты, а именно γ-аминомасляная кислота (ГАМК) и глицин. ГАМКA-рецепторы обнаружены прежде всего в коре больших полушарий и мозжечке, а рецепторы глицина — преимущественно в стволе мозга и в спинном мозге. С точки зрения их генетического происхождения оба рецептора классифицируются как принадлежащие к суперсемейству nAch-рецепторов, для которых характерны четырехсегментная топология и пентамерная стехиометрия субъединиц.
При этом ГАМКA-рецепторы состоят из двух α (α1-6)- и двух β (β1-3)-субъединиц, а также одной γ-, δ-, ε- или π-субъединицы; что касается рецепторов глицина, то это гетеропентамеры из трех α (α1-4)- и двух β (β1)-субъединиц (табл. 1).
2. Воротные механизмы. Для переключения (открывания/закрывания) ГАМКA-рецепторов и рецепторов глицина характерны те же принципы, что и для воротных механизмов каналов nAchR и iGluR. Однако у тех и других каналов различная ионная проницаемость. ГАМКA-рецепторы и рецепторы глицина проявляют высокую селективность в отношении отрицательно заряженных ионов Cl-, следовательно, это медиаторуправляемые хлорные каналы.
Такая избирательность к анионам определяется, по-видимому, свойствами порообразующего М2-сегмента, который по сравнению с аналогичным элементом катионселективного рецепторного канала имеет меньшее число, а также иное размещение положительно заряженных аминокислотных остатков. Влияние ГАМКA-рецепторов и рецепторов глицина на мембранный потенциал зависит от внутриклеточной концентрации Cl-. Если равновесный потенциал для Cl- более отрицателен, чем потенциал покоя, то открываются лигандуправляемые Cl--каналы, что обеспечивает гиперполяризацию постсинаптической мембраны (гиперполяризующее торможение).
Если же равновесный потенциал для Cl- соответствует потенциалу покоя, то при открывании каналов мембранный потенциал не изменяется, однако в результате снижения входного сопротивления возникает тормозный эффект (шунтирующее торможение). В итоге в некоторых ситуациях (например, в период раннего постнатального развития или при патологических состояниях) равновесный потенциал для Cl- может принимать более положительные значения, чем потенциал покоя. В таких условиях активация ли-гандуправляемых Cl--каналов приводит к деполяризации постсинаптической мембраны, а в особых случаях даже к возбуждению постсинаптической клетки (т. е. к генерации потенциалов действия).
3. Фармакология ГАМКA-рецепторов и рецепторов глицина. ГАМКA-рецепторы — это молекулярные мишени для веществ, применяемых в качестве лекарственных средств, а также широко распространенных как наркотики. К таким веществам относятся бензодиазепины (диазепам, клоназепам — известные анксиолитики) и барбитураты (фенобарбитал), используемые как снотворные и седативные средства.
1. Лигандактивируемые ионные каналы. Активация рецепторного ионного канала может осуществляться в результате изменения мембранного потенциала или связывания внеклеточного медиатора (лиганда). Ионные каналы, способные активироваться таким образом, называются лигандактивируемыми каналами или ионотропными рецепторами.
2. Ионные каналы возбуждающих рецепторов. Важнейшие возбуждающие рецепторы — это ионотропные рецепторы глутамата и ионотропные рецепторы ацетилхолина. Они состоят из четырех или пяти субъединиц.
При отсутствии агониста каналы находятся в закрытом состоянии; связывание с агонистом изменяет конформацию участка связывания и его окружения, что приводит к открыванию канала.
3. Ионные каналы тормозных рецепторов. Важнейшие тормозные медиаторы центральной нервной системы — это γ-аминомасляная кислота (ГАМК) и глицин; соответствующие рецепторные каналы — ГАМКA-рецепторы и рецепторы глицина. Их молекулы образованы пятью субъединицами.
- Рекомендуем ознакомиться далее "Мембранный потенциал покоя и потенциалы действия - с точки зрения физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 10.9.2024