
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Ритмическая активность и кодирование информации в нервной системе - с точки зрения физиологии человека
Оглавление статьи:- Возникновение серии потенциалов действия в нервных клетках
- Кодирование информации в нервной системе
- Резюме
- Список использованной литературы
а) Возникновение серии потенциалов действия в нервных клетках. Длительная деполяризация может вызывать в нервных клетках серии ПД. При повышении интенсивности стимула возрастает частота ПД.
1. Ритмическая активность и рефрактерный период. В ответ на кратковременное пропускание стимулирующего тока нервные клетки генерируют одиночный ПД. Что происходит в случае длительной стимуляции, например при поступлении тонических сенсорных потенциалов или суммации ВПСП? Инициация одиночного ПД составляет исключение. Обычно возникают серии импульсов (рис. ниже). Максимальная их частота зависит от абсолютного рефрактерного периода.
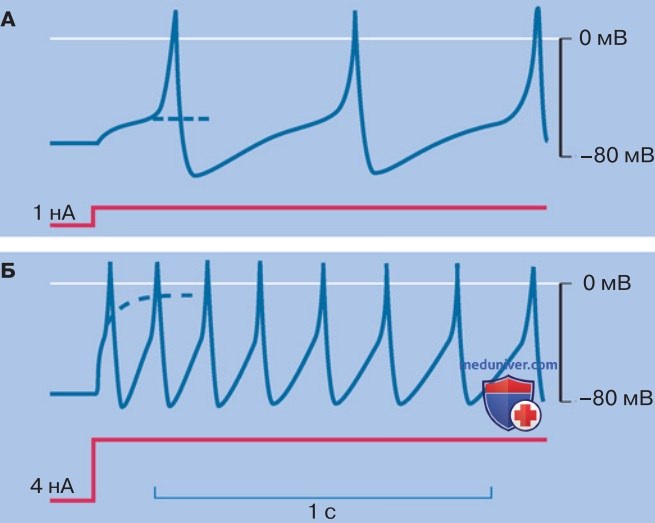
2. Адаптация. Что касается частоты ПД, то она существенно зависит от типа нейрона. Например, в пирамидных клетках коры мозга максимальная частота ПД невелика. Кроме того, эти клетки проявляют выраженную адаптацию, т. е. в условиях постоянной стимуляции частота ПД постепенно снижается. Во время длительной стимуляции наблюдаются ритмические серии ПД, причем промежуток между двумя соседними ПД постепенно нарастает (рис. выше).
Подобное явление объясняется присутствием Са2+-активируемых К+-каналов (КCa) БК-типа (рис. ниже). Во время серии потенциалов действия БК-каналы все более интенсивно активируются входящим Са2+-током, так что амплитуда медленной следовой гиперполяризации, обусловленной БК-каналами, растет. Наряду с активацией БК-каналов существенную роль в адаптационном процессе играет кумулятивная инактивация NaV-каналов.
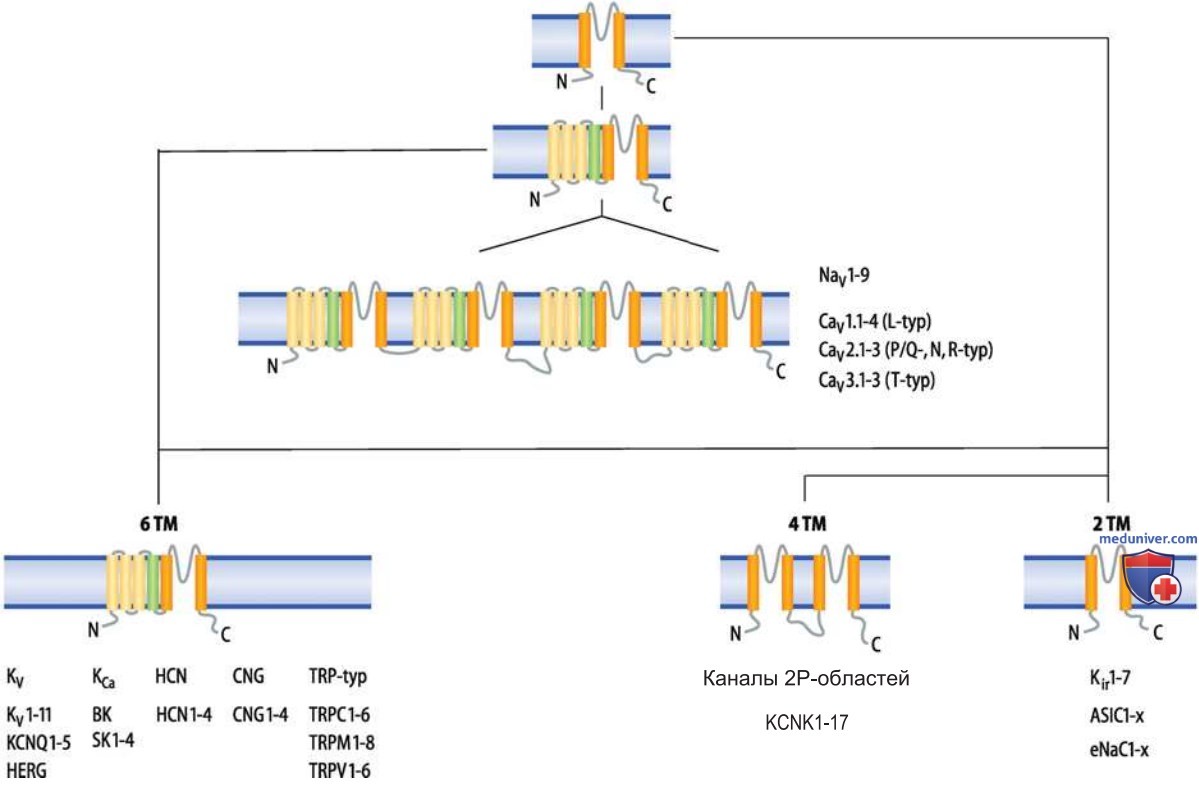
3. Высокочастотные разряды потенциалов действия. Для тормозных интернейронов определенного типа, так называемых быстроразряжающихся, или быстрых (fast-spiking) интернейронов, которые высвобождают ГАМК в качестве медиатора, характерен очень высокий уровень максимальной частоты ПД. При длительной стимуляции генерируются до нескольких сотен ПД в секунду. Причина этого состоит в экспрессии потенциалактивируемых KV3-каналов (рис. выше).
Они отличаются от других типов KV-каналов высоким порогом активации, слабой инактивацией и быстрой кинетикой активации и деактивации. При таких свойствах нетрудно объяснить склонность KV3-каналов к генерации высокочастотных разрядов: вследствие быстрой активации KV3-каналов возрастает скорость их реполяризации и ПД укорачиваются. Из-за быстрой деактивации следовая гиперполяризация длится недолго.
При этом KV3-каналы обеспечивают эффективный восстанавливающий сигнал, благодаря которому NaV-каналы, с одной стороны, выходят из инактивированного состояния, а с другой стороны, сохраняется низкий порог для инициации следующего импульса.
4. Спонтанная ритмическая активность. Некоторые типы нервных клеток (например, мотонейроны или пирамидные клетки коры) в отсутствие входных синаптических сигналов сохраняют стабильный потенциал покоя. Многие нейроны в этой ситуации спонтанно генерируют ПД, например клетки Пуркинье мозжечка и нейроны базальных составляющих ганглиев. Основу спонтанной активности составляют разнообразные молекулярные механизмы.
- Специализированные NaV-каналы после ПД распространяют Na+-ток на соседний участок мембраны, обеспечивая генерацию следующего импульса.
- По окончании ПД инактивирующиеся КV-каналы (например, КV4) сначала восстанавливаются от инактивации, затем снова вступают в фазу инактивации; в итоге возрастает расстояние между двумя соседними ПД.
- Участие каналов, активируемых гиперполяризацией (HCN-каналов).
- Участие СаV-каналов Т-типа (СаV3).
Рассматривая участие HCN- и СаV3-каналов в генерации спонтанной активности нейронов, следует отметить сходство с активностью пейсмекера сердца.
б) Кодирование информации в нервной системе. В нервной системе сигналы перекодируются из аналоговой формы в цифровую; например, градуальные рецепторные потенциалы преобразуются в ПД, соответствующие принципу «все или ничего». Таким образом, ПД можно рассматривать в качестве единиц цифрового нейронного кода, в котором играют роль как частота, так и время возникновения ПД.
1. Потенциал действия как единица цифрового кода. Передача сигнала в нервной системе заключается в проведении ПД от места их генерации (обычно в начальном сегменте аксона — аксонном холмике) через весь аксон к пресинаптическим элементам выходного синапса. Это означает, что ПД опосредуют весь поток информации между нервными клетками. Учитывая, что ПД подчиняются принципу «все или ничего», их можно рассматривать в качестве элементов цифрового сигнала. В кодировании информации играют роль средняя частота и точное время инициации ПД.
Следовательно, здесь имеет место частотное кодирование (кодирование скорости передачи информации) и временное кодирование.
2. Частотное кодирование в нервной системе. Примером может служить регуляция силы сокращения скелетных мышц во время произвольных движений. При низкой частоте разряда α-мотонейронов суперпозиция сократительных реакций подчиненных им двигательных единиц незначительна и вся мышца сокращается слабо. Высокая частота разряда а-мотонейронов обеспечивает выраженную суммацию сократительных реакций двигательных единиц, вплоть до развития тетануса. Следовательно, сила мышечного сокращения регулируется путем частотного кодирования.
3. Временное кодирование в нервной системе. Примером служит детекция источников звука слуховой системой (направленный слух). Нейроны верхней оливы ствола мозга способны сравнивать время возникновения ПД в ипсилатеральных и в контралатеральных входах с точностью до микросекунд. Таким образом, локализация направления, по которому поступает звук, основана на механизме временного кодирования. По-видимому, частотное и временное кодирование осуществляются в коре мозга млекопитающих параллельно.
4. Преобразование аналоговых сигналов в цифровые. В процессе передачи информации в нервной системе аналоговые (градуальные) сигналы должны перекодироваться в цифровую форму (соответствующую принципу «все или ничего»), например преобразование сигналов в мышечных афферентах Ia-типа. При растяжении аннулоспиральных окончаний мышечных веретен наблюдается тоническая деполяризация окончаний, т. е. градуальный рецепторный потенциал. Если деполяризация достигнет порога, в соответствующем афферентном нервном волокне возникнет серия ПД, частота которых возрастает по мере повышения амплитуды деполяризации.
Преобразование аналоговых сигналов в цифровую форму — необходимое условие для осуществления удаленной коммуникации в нервной системе.
в) Резюме. Как правило, длительные стимулы вызывают ритмическую активность нервных клеток. Причем между типами клеток существуют значительные различия. Для пирамидных клеток характерны низкочастотные ПД с выраженной адаптацией, для некоторых ГАМКергических интернейронов, наоборот, ПД с высокой частотой и минимальной адаптацией. Генерация высокочастотных разрядов обусловлена экспрессией КV3-каналов. Спонтанные разряды нейронов возможны даже в отсутствие входных синаптических сигналов.
Механизмы такой спонтанной активности разнообразны, в том числе экспрессия NaV-, KV4-, HCN- и СаV3-каналов. Аналоговые сигналы (например, градуальные рецепторные потенциалы) перекодируются в нервной системе в цифровую форму (ПД, возникающие по принципу «все или ничего»). В кодировании информации играют роль средняя частота ПД (частотное кодирование) и время возникновения ПД (временное кодирование).
Пример частотного кодирования — регуляция силы сокращения скелетной мышцы посредством изменений частоты ПД мотонейронов.
Пример временного кодирования — детекция источников звука слуховой системой ствола мозга (направленный слух).
- Вернуться в раздел "Физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 10.9.2024