
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Транспорт меланосом внутри меланоцитов и к кератиноцитам
а) Транспорт меланосом внутри меланоцитов. Меланосомы транспортируются от места своего происхождения в меланоцитарном прекарионе к кончикам меланоцитарных дендритов. Транспортировка меланосом происходит на микротубулах, расположенных параллельно продольной оси дендрита, и контролируется двумя классами соединенных с микротубулами моторных белков: кинезинами и цитоплазматическими динеинами. Оба двигательных белка действуют как кроткие структурные перекидные «мостики», соединяющие органеллу с микротубулами.
Центробежное антерградное движение органелл опосредуется в основном кинезином, в то время как центростремительное движение контролируется цитоплазматическим динеином. Исследования транспорта меланосом позволяют предположить, что зависящее от микротубул перемещение меланосом происходит в двух направлениях в соответствии с одновременными толчками вперед и назад, исходящими соответственно от кинезина и динеина.
Для меланосом, осуществляющих только центробежное перемещение, двунаправленное движение прекращается после захвата меланосом миозином-Va (кодируется локусом растворения) в богатых актином периферических зонах дендрита.
К белкам, участвующим в транспорте меланосом, относится также Rab27a (кодируется локусом ashen), который опосредует связывание миозина-Va с меланосомами с непосредственным участием другого связующего белка меланофилина (кодируется локусом leaden). При отсутствии миозина-Va меланосомы в кончиках дендритов не скапливаются.
Мутации в любом из перечисленных выше генных продуктов приводят к уменьшению пигментации кожи. Синдром Грисчелли, редкое аутосомно-рецессивное заболевание, при котором наблюдается бледный цвет кожи и волос, является результатом мутации генов миозина-Va, Rab27a и меланофилина. Гены миозина-Va и Rab27a расположены в близком соседстве на хромосоме 15. Поскольку миозин-Va экспрессирован и в мозге, мутации этого гена могут вызывать неврологические аномалии. Rab27a играет также определенную роль в иммунорегуляции, поэтому у лиц с мутациями этого гена наблюдаются нарушения функции иммунной системы. Мутации гена меланофилина приводят только к гипопигментации, характерной для данного синдрома.
б) Транспорт меланосом к кератиноцитам. Транспортировка меланосом от меланоцитов к соседним кератиноцитам является главным этапом нормальной пигментации. Исследования предполагают наличие нескольких путей транспортировки меланосом, включая экзоцитоз, цитофагоцитоз, слияние плазматических мембран и транспортировку мембранными везикулами.
Экзоцитозный способ транспортировки меланосом включает слияние мембраны меланосомы и плазмалеммой меланоцита, высвобождение меланосомы в межклеточное пространство и фагоцитоз окружающими кератиноцитами. Термин цитофагоцитоз указывает на фагоцитоз живой клетки или ее части. Что касается кератиноцитов, они осуществляют цитофагоцитоз кончика меланоцитарного дендрита, который затем сливается с лизосомами внутри кератиноцитов и транспортируется в зону, расположенную над ядром, где фаголизосомные мембраны разрываются, высвобождая меланосомы.
Слияние плазматических мембран кератиноцита и меланоцита образует пространство, через которое меланосомы транспортируются от меланоцита к кератиноциту. Действительно, фотография с высоким разрешением позволяет увидеть присутствие филоподий, тонких, нитевидных, заостренных выступов цитоплазмы у кончиков меланоцитарных дендритов. Перед транспортировкой меланосомы эти филоподии скрепляются и сливаются с плазмалеммой кератиноцита. Четвертый способ транспорта меланосом состоит в отделении наполненных меланосомами везикул, после чего кератиноцитами осуществляется фагоцитоз этих везикул или же происходит их слияние с плазматической мембраной кератиноцитов.
Молекулярные и клеточные механизмы, задействованные в фагоцитозе меланосом, отчасти уже изучены. Установлено, что кератиноциты экспрессируют седьмой трансмембранный сопряженный с G-белком рецептор, который обозначается как активируемый протеазами рецептор 2 типа (PAR-2). PAR-2 активируется, когда сериновые протеазы отщепляют экстрацеллюлярную часть рецептора, открывая новый сегмент, функционирующий как связывающий лиагнт. Активация PAR-2 повышает фагоцитарную активность кератиноцитов.
Примечательно, что УФ-излучение индуцирует активность и экспрессию PAR-2 (это соотносится с ролью УФ в фагоцитозе меланосом). Влияние УФ-излучения на активность и экспрессию PAR-2 более выражено при фототипах кожи II и III, чем при фототипе I. Также в процессе фагоцитоза меланосом кератиноцитами задействован рецептор фактора роста кератиноцитов.
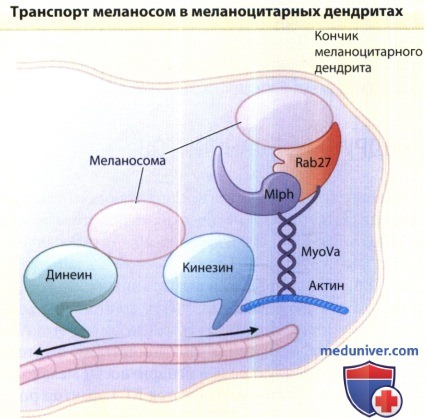
Меланосомы движутся в меланоцитарных дендритах в двух направлениях.
Они прикреплены к микротубулам моторными белками кинезином (антроградное движение) и динеином (ретроградное движение).
У кончика дентрита, меланосомы «застревают» в богатой актином периферии.
Миозин-Va (MyoVa) опосредует связывание меланосом с актином через связующие белки Rab27a и меланофилин (Mlph).
- Рекомендуем далее ознакомиться со статьей "Стимуляция и ингибирование синтеза меланина (регуляция функции меланоцитов)"
Оглавление темы "Меланоциты и меланин.":- Строение и функции меланоцитов
- Меланизация и образование меланосом
- Меланогенные белки влияющие на синтез меланина
- Этапы синтеза меланина
- Строение и функции дендритов меланоцитов
- Транспорт меланосом внутри меланоцитов и к кератиноцитам
- Стимуляция и ингибирование синтеза меланина (регуляция функции меланоцитов)
- Сигнальные пути регулирующие функцию меланоцитов
- Этапы и механизмы загара в ответ на ультрафиолетовое облучение
- Повреждение ДНК клеток при меланогенезе - загаре
- Фотостарение и старение меланоцитов