
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Сигнальные пути регулирующие функцию меланоцитов
Факторы роста, цитокины, гормоны и другие рецепторные лиганды, которые экспрессированы на меланоцитах, осуществляют свою биологическую функцию, взаимодействуя со специфическими рецепторами на своей клеточной поверхности и генерируя сигнальный каскад, вызывающий активацию или ингибицию протеинкиназ и приводящий к различным видам фосфорилирования белков.
В клеточной сигнализации участвуют два типа киназ: сериновые/треониновые киназы и тирозинкиназы, которые по определению фосфорилируют соответственно сериновые и/или треониновые, либо тирозиновые остатки на своих специфических белках-мишенях. В этом разделе рассматриваются основные сигнальные пути, которые влияют на поведение меланоцитов в коже.
а) цАМФ/РКА-зависимый сигнальный путь меланоцитов. Циклический аденозинмонофосфат (цАМФ), идентифицированный одним из первых в качестве вторичного мессенджера, играет ключевую роль в различных биологических функциях, таких как клеточный метаболизм, рост и дифференцировка. Он опосредует также действие a-MHS и был одним из первых распознан в качестве регулятора пигментации у млекопитающих.
Внутриклеточный уровень цАМФ повышается под воздействием мембраносвязанного фермента аденилатциклазы, которая активируется при взаимодействии рецептора и лиганда в рецепторах, спаренных с гуанозинтрифосфат-связывающими белками, такими как MC1R. Уровень цАМФ повышается также под действием таких реагентов, как холераген или изобутилме-тилксантин. Поступление в меланоциты дибутилового аналога цАМФ повышает внутриклеточный уровень цАМФ и индуцирует сигнальный каскад, который приводит к меланогенезу.
цАМФ-зависимая протеинкиназа А (РКА) опосредует большинство биологических действий цАМФ. РКА — это сериновая/треониновая киназа, состоящая из двух регуляторных и двух каталитических субъединиц. В цитозоли она существует в неактивной форме. Связывание цАМФ с регуляторными субъединицами высвобождает каталитические субъединицы, которые активируют фермент. РКА фосфорилирует CREB, который в промоутере MITF связывается с консенсусной последовательностью CRE в ДНК циклического аденозинмонофосфата и вызывает транскрипцию MITF. Повышение уровня цАМФ влияет также и на другие гены-мишени, увеличивая или уменьшая их транскрипцию.
In vitro эффекту РКА можно противодействовать ингибитором протеинкиназы, который выступает в качестве псевдосубстрата для каталитической субъединицы РКА и, таким образом, препятствует фосфорилированию ее эндогенных субстратов.
б) Сигнальный путь меланоцитов с участием протеинкиназы С. Протеинкиназа С (РКС) — сериновая/треониновая киназа — участвует в различных функциях клетки, включая рост, трансформацию и дифференцировку. В качестве неактивного фермента РКС существует в цитоплазме и активируется диацилглицеролом (DAG), компонентом, отщепляющимся от плазмалеммы, когда рецепторы клеточной поверхности взаимодействуют со своими лигандами. DAG высвобождается из плазмалеммы также под действием УФ-излучения. DAG индуцирует перемещение РКС в мембраны, где РКС активируется, вызывая, подобно тирозиназе, фосфорилирование сериновых/треониновых остатков на белках-мишенях. Форболовые эфиры имитируют действие DAG и вначале активируют РКС.
Однако в течение 24 часов все клеточные резервы РКС уничтожаются, и когда на меланоциты воздействуют форболовым эфиром, они уже не способны осуществлять сигнализацию посредством РКС.
Предположение о важной роли РКС в меланогенезе было выдвинуто, исходя из того, что добавление DAG, эндогенного активатора РКС, к культивированным меланоцитам человека приводило к быстрому повышению общего содержания меланина, причем это повышение блокировалось ингибитором РКС. Более того, нанесение DAG на кожу морской свинки увеличивало содержание меланина в эпидермисе.
Экспрессия 12-ти изоформ РКС неодинакова в различных тканях. Считается, что каждая изоформа выполняет определенную биологическую функцию. Меланоциты человека экспрессируют изоформы РКС-α β,-ε, -δ и ς, причем изоформа РКС-β участвует в регулировке активности тирозиназы. ЕТ-1 и гистамин также используют РКС-зависи-мый путь сигнализации (в дополнение к цАМФ-зависимому пути) для своих регуляторных воздействий на функцию меланоцитов.
в) Рецепторы тирозинкиназы. Меланоциты экспрессируют несколько различных рецепторов тирозинкиназы, которые связывают лиганды BMP, bFGF, HCF и c-Kit. Взаимодействие рецептора и лиганда активирует внутриклеточный домен тирозинкиназы, фосфорилирует рецептор и затем активирует серию киназ, называемых MAP-киназами, или же другие внутриклеточные сигнальные молекулы. Через цепную реакцию, которая включает фосфорилирование протеиназ, таких как MITF, сигналы передаются в ядро и активируют, или подавляют транскрипцию генов, участвующих в пролиферации меланоцитов, меланогенезе и/или выживании меланоцитов.
г) β2 и α1-адренергические рецепторы. По данным ряда исследований пути, приводящие к повышению внутри клетки цАМФ, также участвуют в регуляции меланогенеза. Мыши с дефицитом РОМС (РОМС-/-), у которых отсутствует лиганд меланокортина, несмотря на это сохраняют нормальную черную окраску шерсти. При этом с помощью гистологического исследования и электронной парамагнитной резонансной спектрометрии волосяных фолликулов было установлено, что волосяные фолликулы сохраняют нормальную структуру и эумеланиновую пигментацию.
Согласно выдвинутому в этом исследовании положению, либо MC1R сохраняет адекватную базовую активность в отношении индукции пигментации, либо пути, не задействующие меланокортин, также могут индуцировать меланогенез. В самом деле, меланоциты экспрессирут и β2-AR и α1-AR, соответственно. α1-AR взаимодействует с меланоцитарного происхождения норадреналином и повышает уровень DAG, что приводит к индукции меланогенеза по РКС-β-опосредованному механизму. Кератиноциты также вырабатывают адреналин, который соединяется с экспрессируемыми на меланоцитах рецепторами β2-AR, что приводит к повышению уровня цАМФ и синтезу меланина. Поэтому в регуляции меланогенеза могут быть задействованы различные пути, действующие синергично.
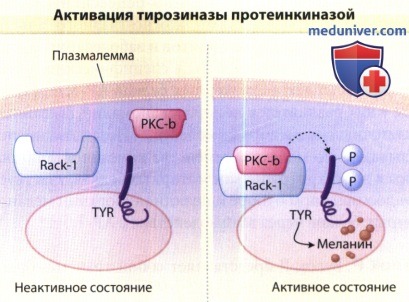
В исходном состоянии отсутствует активация РКС-β и тирозиназа (TYR) не фосфорилируется.
Активированная РКС-β связывает рецепторы для активированной С-киназы-1 (RACK-I), этот комплекс перемещается в меланосому и фосфорилирует сериновые остатки на цитоплазматическом хвосте тирозиназы.
Фосфорилированиетирозиназы активирует фермент, катализирующий биосинтез меланина.
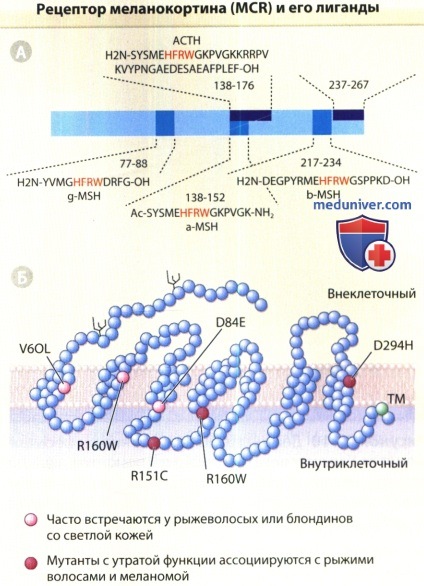
А. Структура прекурсора проопиомеланокортина. Используются стандартные сокращения для аминокислот.
Синтетический аналог суперактивного меланоцитостимулирующего гормона α (α-MSH),
обозначаемый [Nle4 D-Phe7]-a-MSH, модифицирован заменой метионина (М) на норлейцин и L-фенилаланина (F) на D-фенилаланин.
Красным обозначены важные аминокислоты, необходимые для связывания MCR.
Б. Схематическое изображение рецептора MC1R у человека. Каждый из 318 аминокислотных остатков в полипептидной цепи рецептора представлен пустым кружком.
Ответвляющиеся структуры представляют собой N-сцепленные участки гликозилирования.
Указаны мутанты с ослабленной функцией (красные кружки), варианты, распространенные у рыжеволосых и блондинов со светлой кожей (розовые кружки) и консервативный С-терминальный цистеин (зеленый кружок),
возможный участок ацилирования жирной кислоты и прикрепления к плазмалемме.
Ас = ацетилированный; АСТН = адренокортикотропный гормон; NH2 = амидированный; ТМ = трансмембранный домен.
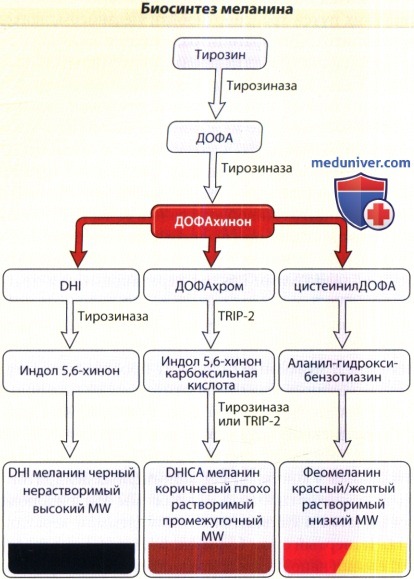
Биосинтез меланина с аминокислотой тирозином, которая преобразуется в L-ДОФА (3,3-дигидроксифенилаланин) на ограничивающем скорость меланогенеза этапе биосинтеза, который катализируется тирозиназой.
L-ДОФА затем превращается в ДОФАхинон посредством того же фермента. После этого образуются DHI (5,6-дигидроксииндол) и DHICA (5,6 дигидроксииндол-2-карбоксильная кислота),
которые производят черный или коричневый эумеланин. Альтернативно, посредством инкорпорации глютатиона или цистеина, ДОФАхинон может образовывать феомеланин.
MW = молекулярный вес; TRP = родственный тирозиназе белок.
- Рекомендуем далее ознакомиться со статьей "Этапы и механизмы загара в ответ на ультрафиолетовое облучение"
Оглавление темы "Меланоциты и меланин.":- Строение и функции меланоцитов
- Меланизация и образование меланосом
- Меланогенные белки влияющие на синтез меланина
- Этапы синтеза меланина
- Строение и функции дендритов меланоцитов
- Транспорт меланосом внутри меланоцитов и к кератиноцитам
- Стимуляция и ингибирование синтеза меланина (регуляция функции меланоцитов)
- Сигнальные пути регулирующие функцию меланоцитов
- Этапы и механизмы загара в ответ на ультрафиолетовое облучение
- Повреждение ДНК клеток при меланогенезе - загаре
- Фотостарение и старение меланоцитов