
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Этапы синтеза меланина
В меланосомах синтезируется два типа меланина: эумеланин и феомеланин. Эумеланин — темный, коричневый или черный, нерастворимый, а феомеланин — светлый, красный или желтый, растворимый и содержит серу. Оба меланина являются индоловыми производными ДОФА и образуются в меланосомах в ходе нескольких этапов окисления. Уровень pH в меланосомах воздействует на активность меланогенных ферментов и влияет на полимеризацию меланина.
Синтез обоих типов меланина проходит через каталитический этап, который определяет его скорость. На этом этапе тирозин окисляется ферментом тирозиназой (которую называют также тирозиноксидаза, монофенол, L-ДОФА: кислород-оксиредуктаза) в L-ДОФА. Этот процесс известен как схема Рэпера-Мэзона.
Преобразование тирозина в L-ДОФА считается основным этапом, ограничивающим скорость меланогенеза, поскольку ингибирование этой реакции блокирует синтез меланина. В реакции синтеза обоих типов меланина L-ДОФА действует в качестве кофактора, а также субстрата тирозиназы. Хотя точные взаимодействия между тирозиназой и ее субстратами полностью не выяснены, кинетические исследования in vitro позволяют предположить, что связывание тирозиназы с тирозином и L-ДОФА опосредуется специальными участками, а связывание с L-ДОФА вызывает конформационные изменения в тирозиназе, которые приводят к повышению аффиности тирозиназы как к тирозину, так и к L-ДОФА.
ДОФА окисляется в ДОФАхинон, ДОФАхинон затем преобразуется в ДОФАхром, а ДОФАхром может трансформироваться в 5,6-дигидроксииндол (DHI) или в 5,6-дигидрокси-2-карбоксильную кислоту (DHICA). Катализатором последней реакции служит фермент ДОФАхром-таутомераза или TRP-2. Уровни коричневого и черного эумеланина соотносятся также как DHI и DHICA, при этом повышение этого соотношения приводит к образованию черного эумеланина, а понижение связано с появлением коричневого эумеланина. ДОФА-хинон может также соединяться с глютатионом или цистеином, в результате чего образуется цистенил-ДОФА, и это соединение становится затем желтым или красным, растворимым веществом с низким молекулярным весом — феомеланином.
Интересно, что тирозиназа катализирует также более отдаленный этап биосинтеза меланина, а именно конверсию DH1 в индол-5,6-хинон. У мышей фермент TRP-1 (называемый также DHlCA-оксидаза) преобразует DHICA в индол-5,6-хинон-карбоксильную кислоту. Однако роль TRP-1 в биосинтезе меланина у человека еще не установлена.
Основной функцией меланина служит обеспечение защиты от вызванного ультрафиолетом повреждения ДНК путем поглощения и рассеивания УФ-излучения (280-400нм). Поэтому поглощение меланином энергии в этом участке электромагнитного спектра максимальное и постепенно уменьшается на протяжении спектра видимого света. Поглощенный меланином ультрафиолет преобразуется в тепло, менее токсичную форму энергии.
Однако проведенные некоторыми специалистами исследования in vitro показывают, что способность меланина действовать в качестве солнцезащитного экрана ограничена, и что меланин, включенный в крем и нанесенный на кожу, поглощает только от 50 до 75% падающего солнечного света. Возможно, конечно, что в меланосомах in vivo меланин, благодаря локализации над ядром, способен обеспечить более высокий уровень защиты.
Промежуточные продукты синтеза меланина, как и сам меланин, могут быть также вредными для клетки, поскольку, в зависимости от молекулярного веса и состояния полимеризации, могут способствовать индуцированному ультрафиолетом (от 320 до 400 нм) повреждению ДНК, вероятнее всего путем генерирования реактивных форм кислорода. Предполагается, что повышенная частота индуцированных ультрафиолетом меланом у светлокожих рыжеволосых лиц объясняется не только пониженной способностью феомеланина осуществлять защиту от вызванного ультрафиолетом повреждения ДНК, но и мутагенностью феомеланина и, вероятно, других промежуточных продуктов синтеза меланина в результате прооксидантных свойств.
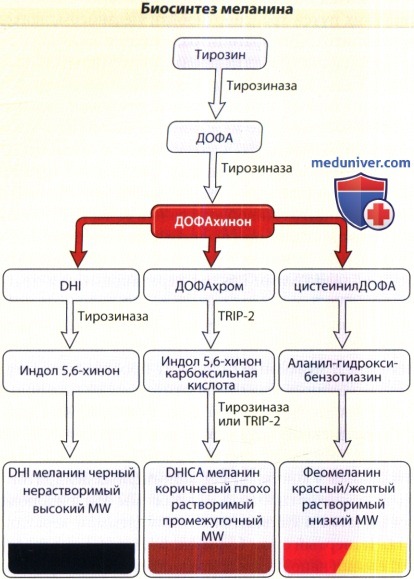
Биосинтез меланина с аминокислотой тирозином, которая преобразуется в L-ДОФА (3,3-дигидроксифенилаланин) на ограничивающем скорость меланогенеза этапе биосинтеза, который катализируется тирозиназой.
L-ДОФА затем превращается в ДОФАхинон посредством того же фермента. После этого образуются DHI (5,6-дигидроксииндол) и DHICA (5,6 дигидроксииндол-2-карбоксильная кислота),
которые производят черный или коричневый эумеланин. Альтернативно, посредством инкорпорации глютатиона или цистеина, ДОФАхинон может образовывать феомеланин.
MW = молекулярный вес; TRP = родственный тирозиназе белок.
- Рекомендуем далее ознакомиться со статьей "Строение и функции дендритов меланоцитов"
Оглавление темы "Меланоциты и меланин.":- Строение и функции меланоцитов
- Меланизация и образование меланосом
- Меланогенные белки влияющие на синтез меланина
- Этапы синтеза меланина
- Строение и функции дендритов меланоцитов
- Транспорт меланосом внутри меланоцитов и к кератиноцитам
- Стимуляция и ингибирование синтеза меланина (регуляция функции меланоцитов)
- Сигнальные пути регулирующие функцию меланоцитов
- Этапы и механизмы загара в ответ на ультрафиолетовое облучение
- Повреждение ДНК клеток при меланогенезе - загаре
- Фотостарение и старение меланоцитов