
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Стимуляция и ингибирование синтеза меланина (регуляция функции меланоцитов)
На поведение меланоцитов в коже в большой степени влияют сигналы от соседних кератиноцитов, а также аутокринные сигналы и факторы окружающей среды, в частности УФ-излучение. Синтез и секреция большинства производимых кератиноцитами факторов усиливается УФ-излучением, но очевидно также, что ультрафиолетовое излучение может непосредственно стимулировать дендритность меланоцитов и производство меланина.
Меланоциты получают как положительные, так и отрицательные паракринные сигналы, которые модулируют пролиферацию и дифференцировку меланоцитов.
I. Стимуляторы меланогенеза:
а) Проопиомеланокортин и производные пептиды. Документально подтверждено, что меланоцитостимулирующий гормон (MSH) и АГТГ являются мощными стимуляторами меланогенеза. Они принадлежат к семейству пептидов, производных перкурсора проопиомеланокортина (POMC), который, кроме гипофиза, синтезируется также эпидермальными кератиноцитами. Интересно, что экспрессия POMC в кератиноцитах индуцируется ультрафиолетом, форболовыми эфирами и интерлейкинами (ИЛ).
У грызунов α-MSH стимулирует меланогенез, причем производство эумеланина преобладает над производством феомеланина, однако системное назначение α-MSH, β-MSH и АКТГ людям увеличивает пигментацию кожи преимущественно на открытых воздействию солнца участках. Однако, при некоторых заболеваниях с аномально высокими уровнями АКТГ, в частности болезни Аддисона или синдроме Нельсона (секретирующая АКТГ аденома гипофиза), наблюдается более генерализованная пигментация кожи.
Сообщалось также, что α-MSH, кроме воздействия на меланогенные белки и синтез эумеланина, стимулирует репарацию вызванного ультрафиолетом повреждения ДНК в меланоцитах, в частности, восстановление димеров пиримидина, а также понижает уровень индуцированной ультрафиолетом перекиси водорода в клетке. Кроме того, установлено, что α-MSH регулирует pH меланосом. Эти данные позволяют предположить, что роль пептидов, производных от РОМС, выходит за рамки стимулирования меланогенеза.
б) Эндотелин-1 (ЕТ-1). ЕТ-1 играет определенную роль в зрелых меланоцитах, индуцируя меланогенез путем активации тирозиназы и повышения показателей TRP-1. ЕТ-1 также вызывает пролиферацию меланоцитов и способствует образованию дендритов. Культивированные кератиноциты синтезируют и секретируют ЕТ-1, а УФ-облучение стимулирует производство ЕТ-1 в кератиноцитах. ЕТ-1 может действовать синергично с другими факторами роста или цитокинами, оказывая, таким образом, дальнейшее влияние на функцию меланоцитов.
ЕТ-1 стимулирует экспрессию уровня MC1R и повышает аффиность MC1R к a-MSH. Подобно α-MSH, ЕТ-1 оказывает фотозащитное действие на меланоциты, усиливая восстановление димеров тимина, уменьшая уровень индуцированной ультрафиолетом перекиси водорода и индуцируя уровень антиапоптозных белков.
в) Фактор стволовых клеток (SF). Как и другие вырабатываемые кератиноцитами факторы, SF индуцируется ультрафиолетовым излучением, и у морских свинок анти-Kit-антитела блокируют загар, вызванный ультрафиолетом. В процессах регуляции индуцированного ультрафиолетом меланогенеза и выживания меланоцитов, SCF может также действовать синергично с другими цитокинами, например, ИЛ-3, ИЛ-6, ИЛ-7, ИЛ-9 и колониестимулирующим фактором гранулоцитов-макрофагов.
г) Медиаторы воспаления. Некоторые медиаторы воспаления могут влиять на пигментацию кожи. PG (простагландины, метаболиты арахидоновой кислоты) и лейкотриены (родственные простагландинам липидные вещества), являясь медиаторами воспалительного ответа, воздействуют и на функцию меланоцитов. Их уровень повышен в загорелой коже, а также при различных воспалительных дерматозах, в том числе атопическом дерматите и псориазе.
Меланоциты человека экспрессируют несколько рецепторов простагландинов, включая рецепторы для PGE2 и PGF2α.Действительно, PGF2α стимулирует образование меланоцитарных дендритов и активирует тирозиназу, а ультрафиолетовое излучение повышает уровень экспрессии рецепторов PG на меланоцитах. Подобным образом лейкотриены В4 и С4 увеличивают синтез меланина и стимулируют пролиферацию и подвижность меланоцитов. Интересно отметить, что меланоциты также способствуют развитию воспалительных реакций кожи, поскольку при стимуляции про-воспалительными цитокинами ИЛ-1 и ФНО-a они синтезируют и выделяют ИЛ-8.
Меланоциты реагируют также на гистамин, выделяемый тучными клетками во время кожного воспаления. Гистамин связывает рецепторы Н1 и Н2, индуцируя дендритность меланоцитов и повышая уровни экспрессии тирозиназы. Эти эффекты уменьшаются, если на меланоциты предварительно воздействовать фамотидином, антагонистом рецептора Н2.
д) Нейротропины. Нейротропины (NT) представляют собой семейство молекул, которые способствуют выживанию нейронов в центральной и периферической нервной системе. К ним относятся NGF, NT3, NT4 и производное мозга — нейротропный фактор. Меланоциты экспрессируют p75NTR, рецептор низкой аффинности, общий для всех NT, а также рецепторы высокой аффинности для NGF (TrkA) и NT3 (TrkC). Синтезируемый в кератиноцитах NGF, экспрессия которого стимулируется УФ-излучением, является хемотаксическим для меланоцитов и индуцирует их дендритность. Как NGF, так и NT3, который экспрессируется фибробластами дермы, увеличивают выживаемость меланоцитов.
В частности, добавление NGF после УФ-облучения увеличивает уровень антиапоптозного белка Bcl2, уменьшая апоптотическую гибель клеток. Таким образом, как и другие производимые кератиноцитами цитокины, NGF помогает сохранить популяцию кожных меланоцитов, которые в противном случае исчезли бы вследствие повреждение ультрафиолетом.
е) Базовый фактор роста фибробластов. Базовый фактор роста фибробластов (bFGF) названный так за способность стимулировать рост фибробластов, был одним из первых идентифицирован как митоген. Он вырабатывается кератиноцитами, но секреторный сигнал при этом отсутствует, поэтому предполагается, что данный фактор воздействует на меланоциты посредством межклеточного контакта. Фактор bFGF связывает трансмембранные рецепторы тирозинкиназы и индуцирует ее митогенное действие в присутствии факторов, повышающих уровни цАМФ. Как и в случае других производимых кератиноцитами цитокинов, экспрессия bFGF стимулируется в ответ на УФ-облучение.
Фактор роста кератиноцитов, еще один член семейства белков FGF, способствует передаче меланосом от меланоцитов в кератиноциты.
ж) Окись азота. Окись азота (NO)—растворимый свободный радикал, оказывающий плеотропное биорегуляторное воздействие на различные клетки и ткани. Меланоциты и кератиноциты вырабатывают NO в ответ на воспалительные цитокины, а производство NO в кератиноцитах индуцируется УФ-излучением. NO увеличивает активность тирозиназы и меланогенез и является, таким образом, аутокринной, а также паракринной молекулой, влияющей на функционирование меланоцитов в коже.
з) Катехоламины. Катехоламины — это группа сигнальных молекул, первично функционирующих как нейротрансмиттеры и как эндокринные гормоны. Катехоламины связываются либо с α1-адренергическими рецепторами (AR), либо с β2-AR и могут индуцировать меланогенез через РКС-β или сАМР-зависимые сигнальные пути.
II. Ингибиторы меланогенеза. В многочисленных сообщениях высказывалось предположение о существовании эндогенных ингибиторов меланогенеза, но пока идентифицировано лишь несколько специфических молекул. К одной из групп ингибиторов относятся сфинголипиды, которые принадлежат к классу мембраносвязанных липидов и действуют в качестве передатчиков сигнала. Было показано, что сфинголипиды ослабляют меланогенез как минимум частично, усиливая распад MITF посредством убиквитина. Еще один ингибитор меланогенеза, ВМР-4, подавляет экспрессию тирозиназы в меланоцитах, частично также воздействуя на MITF.
Интересно, что физиологические дозы УФ-облучения, мощного стимулятора меланогенеза, уменьшают экспрессию рецепторов BMP на меланоцитах, устраняя при этом его ингибирующее действие в процессе вызываемого УФ-облучением загара. Трансгенные мыши с избыточной экспрессией ноггина, физиологического антагониста BMP, имеют более темный цвет шкурки, чем дикие мыши, а в их волосках повышено соотношение эумеланина к феомеланину.
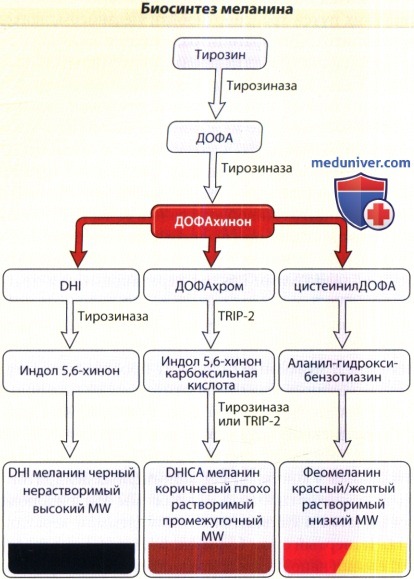
Биосинтез меланина с аминокислотой тирозином, которая преобразуется в L-ДОФА (3,3-дигидроксифенилаланин) на ограничивающем скорость меланогенеза этапе биосинтеза, который катализируется тирозиназой.
L-ДОФА затем превращается в ДОФАхинон посредством того же фермента. После этого образуются DHI (5,6-дигидроксииндол) и DHICA (5,6 дигидроксииндол-2-карбоксильная кислота),
которые производят черный или коричневый эумеланин. Альтернативно, посредством инкорпорации глютатиона или цистеина, ДОФАхинон может образовывать феомеланин.
MW = молекулярный вес; TRP = родственный тирозиназе белок.
- Рекомендуем далее ознакомиться со статьей "Сигнальные пути регулирующие функцию меланоцитов"
Оглавление темы "Меланоциты и меланин.":- Строение и функции меланоцитов
- Меланизация и образование меланосом
- Меланогенные белки влияющие на синтез меланина
- Этапы синтеза меланина
- Строение и функции дендритов меланоцитов
- Транспорт меланосом внутри меланоцитов и к кератиноцитам
- Стимуляция и ингибирование синтеза меланина (регуляция функции меланоцитов)
- Сигнальные пути регулирующие функцию меланоцитов
- Этапы и механизмы загара в ответ на ультрафиолетовое облучение
- Повреждение ДНК клеток при меланогенезе - загаре
- Фотостарение и старение меланоцитов