
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Обмен витамина D и его функции
Основная функция жирорастворимого витамина D — поддержание нормальных уровней кальция и фосфора в крови для обеспечения метаболических функций, минерализации костной ткани и нервно-мышечной передачи. Отсутствие витамина D приводит к заболеваниям костей, например рахиту (у детей с неполным заращением эпифизов) и остеомаляции (у взрослых), а также гипокальциемической тетании.
Последнее заболевание проявляется судорожными припадками из-за недостаточной концентрации во внеклеточной жидкости ионизированного кальция, необходимого для нормального нервного возбуждения и расслабления мышц. В конце XIX в. наблюдалась практически эпидемия рахита в крупных городах Европы и бедных районах Нью-Йорка и Бостона.
Хотя антирахитические свойства рыбьего жира были признаны еще в начале того века, понадобилось почти 100 лет, чтобы его стали использовать в медицинской практике в качестве эффективного профилактического средства (это усложнялось тем, что рыбий жир, употребляемый в рыбацких деревнях Северной Европы, Скандинавии и Исландии, представлял собой темную, плохо пахнущую жидкость). Помимо регуляции гомеостаза кальция и фосфора витамин D имеет и неклассические эффекты (действует и на нескелетные ткани).
Метаболизм витамина D. Основной источник витамина D у людей — эндогенный синтез в коже путем фотохимических преобразований его предшественника (7-дегидрохолестерина) под действием энергии солнечных или искусственных УФ-лучей с длиной волны от 290 до 315 нм (УФ-В-излучение). При облучении 7-дегидрохолестерина образуется холекалъциферол, известный как витамин D3 (в данной книге для упрощения использован термин «витамин D»). В обычных условиях после воздействия солнечных лучей из 7-дегидрохолестерина в коже синтезируется 90% необходимого витамина D.
Однако для афроамериканцев характерен меньший его синтез (10%) из-за меланина. Небольшая часть витамина D поступает с пищей (например, при употреблении глубоководной рыбы, растений и зерновых). В растительных источниках он представлен в форме предшественника (эргостерола), который преобразуется в организме в витамин D.
Метаболизм витамина D в общих чертах можно описать следующим образом:
1. Фотохимический синтез витамина D из 7-дегидрохолестерина в коже и его всасывание из пищи в кишечнике.
2. Связывание с a1-глобулином плазмы (витамин D-связывающий белок) и транспорт в печень.
3. Преобразование в 25-гидровитамин D [25(OH)D] с помощью 25-гидроксилаз (включая CYP27A1 и другие CYP) в печени.
4. Преобразование 25(OH)D а1-гидроксилазой в почках в 1,25-дигидроксивитаминО [1a,25(OH)2D3], или, упрощенно, 1,25(OH)2D (биологически активную форму витамина D).
Синтез 1,25(OH)2D в почках регулируется 3 основными механизмами:
(1) гипокальциемия стимулирует секрецию РТН, который, в свою очередь, увеличивает преобразование 25(OH)D в 1,25(OH)2D, активируя а1-гидроксилазу;
(2) гипофосфатемия непосредственно активирует а1-гидроксилазу, увеличивая образование 1,25(OH)2D;
(3) по механизму обратной связи повышение уровня 1,25(OH)2D приводит к уменьшению синтеза этого метаболита, ингибируя активность а1-гидроксилазы.
Механизм действия витамина D. 1,25(OH)2D лучше всего рассматривать как стероидный гормон. 1,25(OH)2D взаимодействует с высокоаффинным рецептором витамина D, который связан с RXR. Эти гетеродимерные комплексы связываются с ответными элементами витамина D, расположенными в промоторе генов-мишеней витамина D. Рецепторы для 1,25(OH)2D присутствуют в большинстве клеток организма и опосредуют сигналы, регулирующие уровни кальция и фосфора в плазме, действуя на тонкий кишечник, кости и почки.
Помимо действия на костный гомеостаз витамин D также обладает иммуномодулирующим и антипролиферативным эффектами. Есть предположение, что 1,25(OH)2D может действовать через негенетические механизмы, т.е. без транскрипции генов-мишеней. Негенетическим механизмом может быть связывание 1,25(OH)2D с мембранным рецептором витамина D, что приводит к активации протеинкиназы С и открытию кальциевых каналов.
Воздействие на гомеостаз кальция и фосфора. Основными воздействиями 1,25(OH)2D на гомеостаз кальция и фосфора являются:
- стимуляция всасывания кальция в кишечнике. l,25(OH)2D стимулирует абсорбцию кальция в двенадцатиперстной кишке путем взаимодействия с ядерными рецепторами витамина D и формирования комплекса с RXR. Этот комплекс связывается с ответными элементами витамина D и активирует транскрипцию рецептора транзиторного потенциала по ванилло-иду (TRPV) 6, который кодирует важные кальциевые каналы;
- стимуляция процесса реабсорбции кальция в почках. 1,25(OH)2D увеличивает приток кальция в дистальные почечные канальцы путем повышения экспрессии TRPV5, другого рецептора семейства ваниллоидов. Экспрессия TRPV5 также регулируется РТН в ответ на гипокальциемию;
- взаимодействие с РТН при регуляции уровня кальция в крови. Витамин D поддерживает уровень кальция и фосфора в плазме на высоком уровне. Паращитовидные железы играют ключевую роль в регуляции внеклеточной концентрации кальция. Эти железы имеют кальциевый рецептор, чувствительный даже к небольшим изменениям концентрации кальция в крови.
Помимо уже описанного влияния на всасывание кальция в кишечнике и почках l,25(OH)2D и РТН увеличивают экспрессию лиганда RANK (RANKL) остеобластами. RANKL связывается со своим рецептором RANK на преостеокластах, инициируя дифференцировку этих клеток в зрелые остеокласты. С помощью секреции соляной кислоты и активации протеаз, таких как катепсин К, остеокласты разрушают кость и высвобождают кальций и фосфор в кровяное русло;
- минерализация костной ткани. Витамин D способствует минерализации остеоидного матрикса и эпифизарного хряща при формировании как плоских, так и длинных костей скелета. Он стимулирует синтез остеобластами кальцийсвязывающего белка остеокальцина, участвующего в отложении кальция при развитии костей.
Плоские кости формируются путем внутримембранозной оссификации, когда мезенхимальные клетки дифференцируются непосредственно в остеобласты и синтезируют коллагеновый остеоидный матрикс, на котором откладывается кальций. Длинные кости формируются путем энхондральной оссификации. Растущий хрящ эпифизарных пластин временно минерализуется, затем постепенно рассасывается и замещается остеоидным матриксом, который минерализуется уже с образованием кости.
Дефицит витамина D вызывает гипокальциемию. В результате повышается синтез РТН, что активирует почечную а1-гидроксилазу, увеличивающую количество активного витамина D и усвоение кальция. Кроме этого, синтез РТН усиливает резорбцию кальция из костей остеокластами, снижает экскрецию кальция и повышает экскрецию фосфатов почками.
Фактор роста фибробластов 23, продуцируемый костями, является представителем группы фосфатонинов, которые блокируют абсорбцию фосфатов в кишечнике и реабсорбцию их в почках, в результате увеличивается выведение фосфата с мочой. Нормальный уровень кальция в сыворотке крови может восстановиться, но гипофосфатемия сохраняется и нарушается минерализация костей. Повышенный синтез фактора роста фибробластов 23 может привести к опухоль-индуцированной остеомаляции и некоторым формам рахитов, ассоциированных с гипофосфатемией.
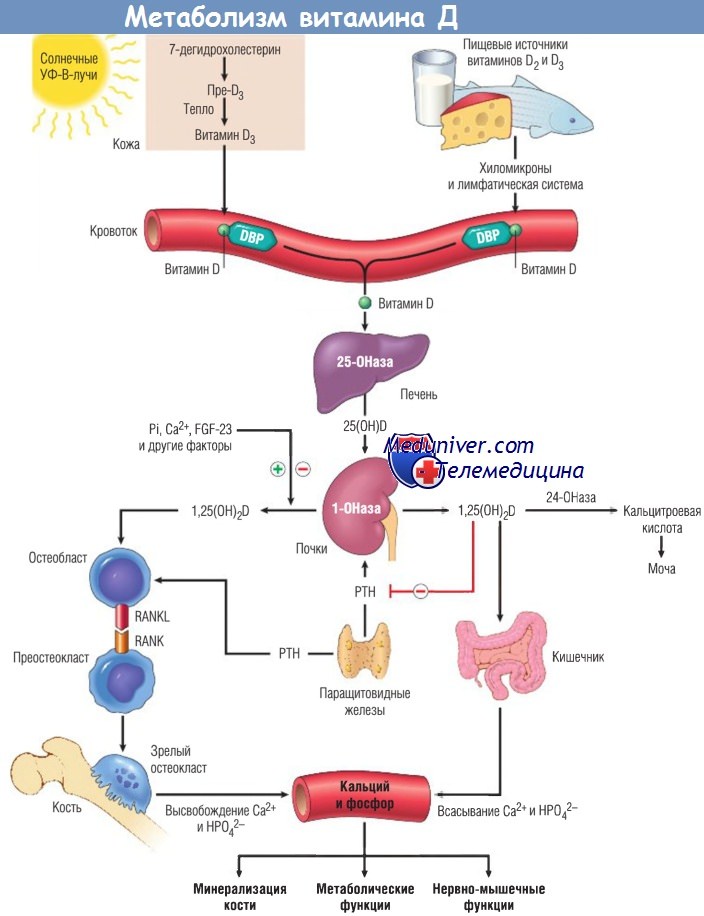
В печени он превращается в 25(OH)D (25-гидроксивитамин D), а в почках — в 1,25(OH)2D (1,25-дигидроксивитамин D), активную форму витамина D.
1,25(OH)2D стимулирует экспрессию лиганда RANK (RANKL) — регулятора созревания и функций остеокластов — на остеобластах, и увеличивает всасывание кальция и фосфора в кишечнике.
DBP — витамин D-связывающий белок (а1-глобулин); FGF — фактор роста фибробластов;
Pi — фосфатидилинозитол; РТН — паратиреоидный гормон; УФ — ультрафиолетовый.
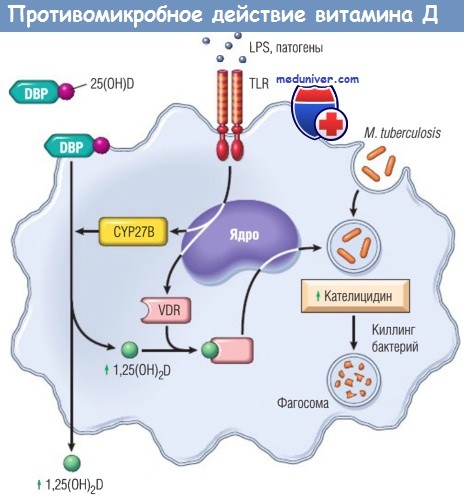
Патогены и липополисахариды (LPS) стимулируют Toll-подобные рецепторы (TLR) макрофагов, вызывая транскрипцию рецептора витамина D (VDR) и повышение активности CYP27B в митохондриях.
Это приводит к образованию 1,25(ОH)2D, который стимулирует синтез кателицидина, антимикробного пептида, особенно активного в отношении М. tuberculosis.
1,25(ОH)2D — 1,25-дигидроксивитамин D; 25(ОH)D — 25-гидроксивитамин D;
CYP27B — изоформа цитохрома Р450; DBP — витамин D-связывающий белок.
- Рекомендуем ознакомиться со следующей статьей "Признаки дефицита витамина D и отравления им"
Оглавление темы "Патофизиология питания":- Обмен витамина А и его функции
- Признаки дефицита витамина А и отравления им
- Обмен витамина D и его функции
- Признаки дефицита витамина D и отравления им
- Обмен витамина С и его функции
- Признаки дефицита витамина С и отравления им
- Механизмы развития (патогенез) ожирения
- Последствия ожирения и почему надо худеть?
- Механизмы влияния питания на развитие опухоли
- Механизмы влияния питания на развитие атеросклероза