
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Эндокринная функция половых желез. Эмбриогенез половой системы
Эндокринная функция половых желез у разных видов животных имеет особый характер. У некоторых беспозвоночных и насекомых выпадение эндокринной функции или пересадка половых желез не оказывает значительного влияния на их рост, на развитие вторичных половых признаков и половое поведение. После удаления зачатков мужских половых желез у гусеницы развитие ее и превращение в самца-бабочку совершаются нормально, причем не утрачиваются даже инстинктивные реакции по отношению к самкам. У многих беспозвоночных кастрация также не оказывает влияния па вторичные половые признаки. У планарий и оболочников половые железы после удаления регенерируют.
Однако у других беспозвоночных, например у ракообразных и кольчатых червей, удаление половых желез прекращает гормональное влияние на развитие вторичных половых признаков. Так, если у самца-краба разрушить половые железы, то полностью исчезают вторичные половые признаки. Это впервые было описано Giard. По данным Smith, при заражении самцов-крабов особым паразитом сакулиной наступает разрушение половых желез (паразитарная кастрация), вследствие чего исчезают мужские половые признаки и самцы по типу приближаются к самкам. У рыб, амфибий и рептилий наличие вторичных половых признаков находится в полной зависимости от гормональной функции половых желез. Выпадение гормонального влияния ведет к недоразвитию или исчезновению вторичных половых признаков и инстинктивных реакций (Я. Д. Киршенблат).
Влияние гормональной функции половых желез на вторичные половые признаки разных птиц проявляется по-разному. У чаек и куликов брачный наряд оперения целиком зависит от гормона семенников, у петуха, курицы, утки оперение и поведение также зависят от полового гормона. У птиц и рептилий среди половых гормонов отсутствует гормон желтого тела.
У северных оленей развитие рогов не зависит от половых гормонов. У млекопитающих эндокринная функция половых желез выражена наиболее ярко, и вместе с тем весьма сложно.
Первичные зародышевые половые клетки у некоторых видов животных появляются вначале во внезародышевой эндодерме желточного мешка.
В человеческом зародыше размером 5—7 мм половые клетки обнаруживаются в первичных половых гребнях, вдоль задней стенки первичной кишки. Они расположены вдоль средней линии брюшной поверхности мезонефрона. Зародышевый эпителий половых гребней разрастается, образуя первичные половые тяжи, проникающие внутрь расположенной под ними мезенхимы. У зародыша размером 14—16 мм первичные половые железы у обоих полов одинаковы. Дифференциация половых желез в яичники или в семенники зависит от преобладания коркового или мозгового компонента. В половой железе, из которой развивается яичник, зародышевый эпителий образует вторичные половые тяжи. После того как они развились, мозговой компонент, включающий первичные половые тяжи, регрессирует и в дальнейшем развивается только корковый слой.
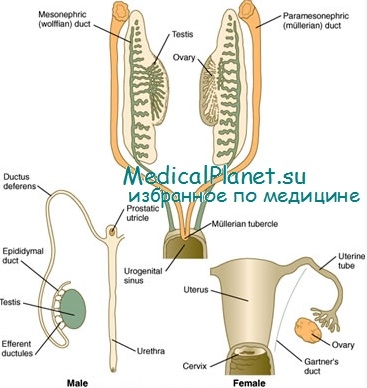
Мозговой или ретикулярный слой яичника содержит рассеянные небольшие сети канальцев и незначительные скопления клеток Лейдига как остатки мозгового компонента. Если начинают развиваться семенники, регрессирует корковый компонент. Канальцевые структуры и межканальцевые клетки, являющиеся основой для развития клеток Лейдига, возникают из мозгового компонента (Wilkins). В дальнейшем у зародыша размером 10 мм проток Мюллера на каждой стороне происходит из мочеполового тяжа и появляется на 6-й неделе. Вольфов проток развивается из верхнего отдела мезонефрических протоков и канальцев и хорошо дифференцируется на 5-й неделе, когда зародыш достигает 23 мм. На 8-й неделе обе группы протоков хорошо развиты. В течение 3-го месяца мюллеровы протоки развиваются по женскому типу, образуя верхнюю часть влагалища, матку и фаллопиевы трубы, а вольфовы протоки исчезают, за исключением зародышевых остатков, которые могут оставаться как элоофорон, пароофорон и соответствующий проток. У лиц мужского пола вольфовы протоки, семявыносящий проток и семенные пузырьки развиваются в придатки яичника, а мюллеровы протоки остаются как остатки придатка яичника.
Green и др. считают, что дифференциация половых протоков контролируется женскими или мужскими гормонами, образуемыми эмбриональными половыми железами.
Wiesmer выдвинул гипотезу автономной тенденции к развитию по женскому типу, если этому не противопоставлены мужские гормоны. Jost удалось кастрировать эмбрион кролика еще до начала какой-либо дифференциации пола. Raymond, Wells произвели такие же опыты на мышах и крысах. Опыты показали, что семенник зародыша осуществляет свое маскулинизирующее действие в короткий критический период. Кастрация, произведенная позже этой критической фазы, дает более выраженную мужскую форму половой системы. Если .кастрация откладывается до срока, когда дифференциация по мужскому типу уже фиксирована, хотя и не полностью, нарушения, полового развития не происходит. Раннее удаление семенника зародыша на одной стороне ведет к развитию фаллопиевой трубы и сохранению более или менее полного рога матки только на кастрированной стороне.
Эти опыты указывают, что при отсутствии зародышевых половых желез мюллеровы протоки развиваются по женскому типу.
Наблюдение Jost и др., установивших, что мюллеров проток остается и продолжает расти при культивировании вне организма, доказывает, что феминизация зародыша с удаленными половыми железами не зависит от материнских или внеполовых феминизирующих веществ. Одностороннее удаление семенника указывает, что секреция распространяется вдоль полового тракта. При инъекции тестостерона плодам женского пола происходит развитие вольфовых протоков и наружных половых органов по мужскому типу.
Обезглавливание эмбриона и удаление гипофиза вызывают уменьшение клеток Лейдига и препятствуют развитию наружных половых органов по мужскому типу. Развитие семенника в поздней стадии вызывается гонадотропными гормонами. Различие 'между наружными половыми органами мужчин и женщин зависит от степени развития полового органа и срастания по средней линии губо-мошоночных складок и других структур. У особей женского пола половой бугорок перестает расти и по сравнению с увеличивающимся ростом тела становится меньше. У особей мужского пола половой бугорок зародыша, развиваясь, образует половой член, губо-мошоночные складки срастаются по средней линии к основанию полового члена, мочевая бороздка закрывается и образуется мошонка и задняя промежуточная часть уретры. К 14-й неделе ткани развиваются и срастаются на нижней поверхности полового члена по средней линии, образуя уретру :и кавернозное тело. У здоровых лиц женского пола губо-мошоно'чные складки не срастаются и мочеполовая борозда остается открытой, образуя половую щель. На 5-м месяце мочеполовая борозда образует глубокое преддверие, куда открывается отверстие уретры и влагалище.
У самцов по мере развития семенники опускаются в мошонку.
Половые различия у млекопитающих определяются следующими особенностями: 1) образование семенников или яичников; 2) образование вспомогательных органов размножения. У самок развиваются матка, яйцевод, влагалище и молочные железы, а у самцов — семенные пузырьки, предстательная железа и половой член. Среди других характерных признаков следует считать и определенные размеры скелета. У женщин таз имеет больший размер, у них также иное распределение жира, рост волос, характер голоса.
- Читать далее "Обследование больной в гинекологии."
Оглавление темы "Гормоны надпочечников":- Гипоальдостеронизм - альдостеронопения. Гиперфункция коры надпочечников
- Первичный гиперальдостеронизм: причины, клиника и диагностика
- Онтогенез мозговой части надпочечников. Хромаффинная ткань
- Гистология и гормоны мозговой части надпочечников
- Иннервация адреналовой ткани надпочечников
- Влияние адреналина и норадреналина на сердечно-сосудистую систему
- Влияние адреналина и норадреналина на обмен веществ
- Гиперфункция мозговой части надпочечников
- Феохромоцитома: частота и патогенез
- Эндокринная функция половых желез. Эмбриогенез половой системы