
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Расположение транспортеров в эпителиальных клетках - с точки зрения физиологии человека
Оглавление статьи:- Вход Na+ через натриевые каналы
- Транспорт глюкозы и аминокислот
- Секреция и абсорбция ионов Cl- посредством Na+, К+, 2Cl--симпорта
- Секреция K+ во внутреннем ухе
- Абсорбция и секреция HCO3-
- Секреция внутриглазной жидкости
- Коротко
- Список использованной литературы
а) Вход Na+ через натриевые каналы. Электрогенный вход Na+ и выход К+ осуществляется через каналы, встроенные в апикальную мембрану клеток дистальных сегментов эпителия.
1. Na+/К+-АТФаза всегда находится в базолатеральной мембране. Для клеток большинства эпителиальных тканей характерно определенное расположение транспортеров: Na+/К+-АТФаза локализуется базолатерально, а другие транспортеры распределены в доменах плазматической мембраны асимметрично. Далее будет рассмотрено типичное местонахождение переносчиков.
2. Проникновение Na+ в клетки эпителия дистальных сегментов трубкообразных органов. В клетки эпителия кишечника, почечных канальцев, потовых и слюнных желез Na+ входит через эпителиальные Иа+-каналы (ЕХаС) апикальной мембраны (рис. 1А). Их избирательный блокатор — диуретик амилорид. В результате входа Иа+ апикальная мембрана деполяризуется.
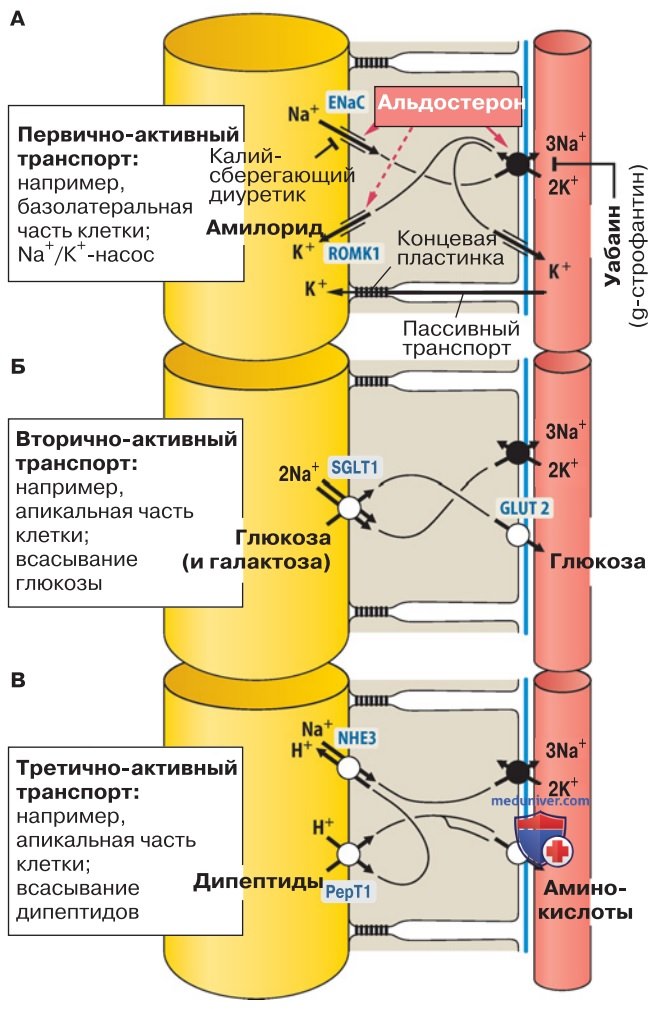
Поскольку мембранный потенциал базолатерального домена не изменился, возникает трансэпителиальная разность потенциалов до -60 мВ, причем отрицательно заряжена поверхность эпителия, обращенная к полости органа. Благодаря этому потенциалу из клеток в полость секретируется K+.
3. Реабсорбция Na+ из альвеол в ткань легких. Отсутствие жидкости в альвеолах необходимо для осуществления газообмена в легких. При этом важна функция ЕХаС, через которые в интерстициальное пространство уходят наиболее распространенные катионы альвеолярной жидкости. Вслед за ними всасываются Cl-, другие растворенные вещества и вода.
б) Транспорт глюкозы и аминокислот. Глюкоза и аминокислоты поступают в эпителиальные клетки проксимальных сегментов с помощью симпортеров апикальной мембраны.
1. Всасывание питательных веществ в виде мономеров. Углеводы всасываются в виде моносахаридов, белки — преимущественно в виде аминокислот или олигопептидов. Транспортеры располагаются главным образом в проксимальных сегментах кишечника и почечных канальцев (рис. 1Б).
2. Моносахариды. Для Na+ и глюкозы существует два вида переносчиков (см. табл. А1).
- Переносчики типа SGLT2 начального (извитого) проксимального почечного канальца обладают низким сродством к глюкозе (перенос в соотношении 1 : 1).
- Переносчики типа SGLT1 прямого участка проксимального почечного канальца (pars recta) и в тонкой кишке имеют более высокое сродство к глюкозе (перенос в соотношении 2:1) и могут работать при очень низкой ее концентрации в полости органа. Наряду с глюкозой переносчик SGLT1 акцептирует галактозу.
Фруктозу переносит через апикальную мембрану только унипортер GLUT5. Выход глюкозы, фруктозы и галактозы через базолатеральную мембрану осуществляется с помощью другого унипортера, GLUT2.
3. Аминокислоты (AS) поступают в клетку с помощью многочисленных транспортеров, в том числе переносчиков кислых, нейтральных и щелочных AS. В большинстве своем это симпортеры, переносящие AS вместе с Na+ (см. табл. А1). В апикальной мембране третично-активные симпортеры поглощают совместно с Н+ часть AS в виде ди- или трипептидов (рис. 1В); в клетке пептиды гидролизуются до AS.
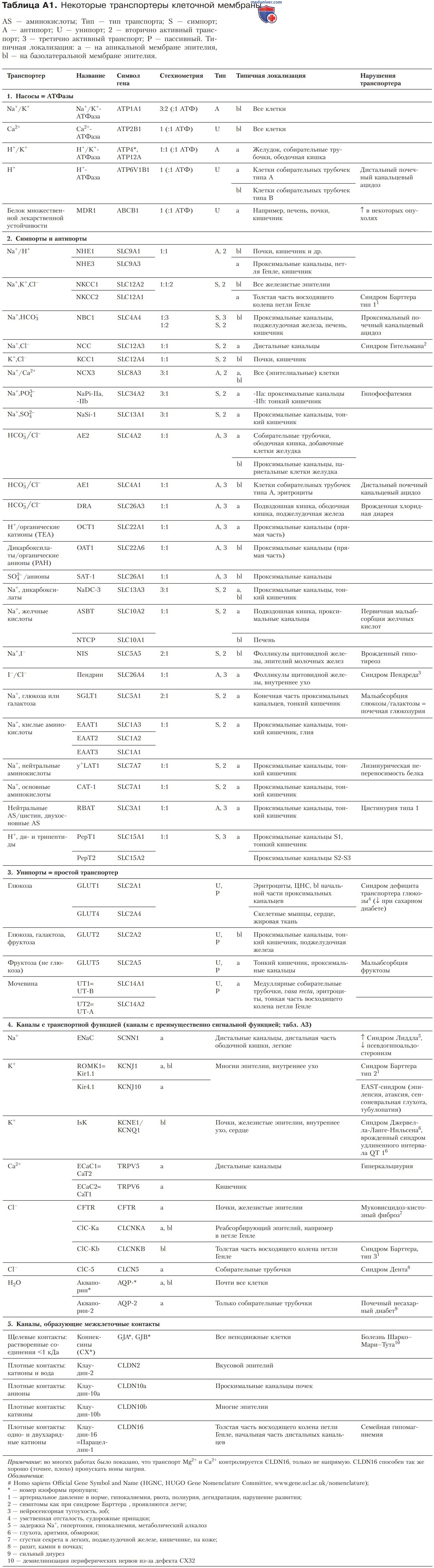
в) Секреция и абсорбция ионов Cl- посредством Na+, К+, 2Cl--симпорта. Секреция Cl- через апикальную мембрану осуществляется благодаря хлорным каналам, а через базолатеральную мембрану — с помощью Na+,К+,2Cl--переносчиков; при абсорбции Cl- соответствующие транспортеры располагаются зеркально.
1. Секреция Cl-. Секреция Cl- запускает выход воды и растворенных веществ (рис. 2). Этот фундаментальный механизм секреторного эпителия действует во всех отделах желудочно-кишечного тракта, протоках от ацинусов желез внешней секреции, дыхательных путях и многих других органах (кроме почек).
Через базолатеральную мембрану Cl- входит в клетку против электрохимического градиента с помощью Na+,К+,2Cl--симпортера (NKCC1) в результате вторично-активного транспорта. Сим-портер NKCC1 блокируют диуретики фуросемид и буметанид. В апикальной части Cl- выходит в полость органа через Ch-каналы CFTR (cystic fibrosis transmembrane conductance regulator).
2. Реабсорбция Cl- осуществляется в утолщенной восходящей части петли Генле. В почках это параклеточный процесс либо частично опосредуемый HCO3-/Cl--антипортером (см. далее). Транспортеры аналогичны тем, которые действуют в секреторной части эпителия, но расположены они зеркально: Na+,K+,2C1--переносчики (NKCC2), блокируемые фуросемидом и буметанидом, находятся в апикальной мембране, а Cl--каналы (CIC-Kb) — в базолатеральной мембране.
P.S. Синдром Бартера:
- Этиология и патогенез. Уже в младенческом возрасте возникают такие симптомы, как гипокалиемия, рвота, полиурия, обезвоживание и нарушение роста, не сопровождающиеся повышением кровяного давления. Причиной заболевания служит мутация гена Nа+,K+,2Cl--симпортера (NKCC2) в восходящей утолщенной части петли Генле (синдром Бартера 1-го типа; табл. А1).
Аналогичные симптомы характерны также при нарушении К+-канала ROMK1 (синдром Бартера 2-го типа) или хлорного Cl--канала C1C-Kb (синдром Бартера 3-го типа) (рис. 2Б). Всасывание NaCl может происходить только при условии, что способны функционировать все три транспортера.
- Псевдосиндром Бартера. Мишенью часто используемого диуретика фуросемида служит сим-портер NKCC2. Блокируя NKCC2, фуросемид вызывает усиленное выделение NaC1 и воды через почки. В связи с этим последствия продолжительного приема фуросемида получили название псевдосиндрома Бартера.
P.S. Синдром Гительмана. Симптомокомплекс аналогичен тому, который наблюдается при синдроме Бартера, но в более стертой форме. Те же симптомы несколько ослаблены. В этом случае в дистальном канальце нарушается поступление Na+Cl- через симпортер NCC апикальной мембраны.
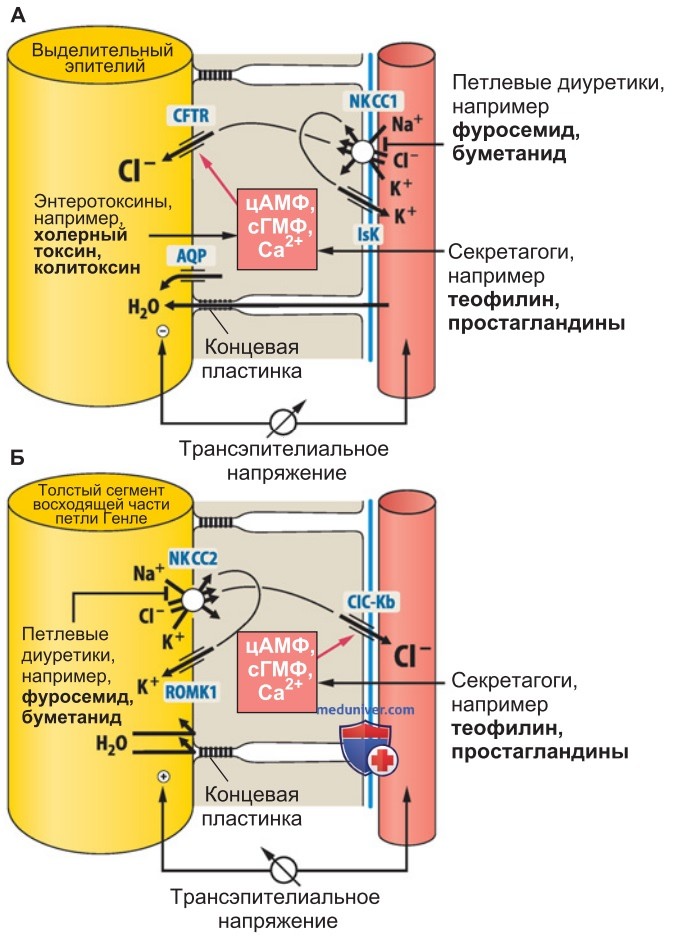
г) Секреция K+ во внутреннем ухе. Чтобы человек нормально слышал, в эндолимфе его внутреннего уха должна быть высокая концентрация K+; диуретик фуросемид, усиливающий секрецию K+ в дистальной части почечных канальцев, может вызвать временную глухоту.
1. Эпителий сосудистой полоски (stria vascularis). Для преобразования акустических сигналов в нервные импульсы необходимы следующие условия: высокий трансэпителиальный потенциал в перепончатом канале улитки (эндокохлеарный потенциал +80 мВ) и высокая концентрация K+ в эндолимфе (150 ммоль/л). Оба фактора обеспечиваются клетками эпителия сосудистой полоски (рис. 3):
- K+-каналы Kir4.1 апикальной мембраны базальных клеток создают высокий эндокохлеарный потенциал;
- маргинальные клетки (составляющие основную часть сосудистой полоски) секретируют K+ в эндолимфу с помощью Nа+,K+,2Cl--симпортера (NKCC1) базолатеральной мембраны и K+-каналов IsK (KCNQ1/KCNE1) апикальной мембраны (рис. 3, справа).
2. Нарушение секреции K+. Диуретик фуросемид может ингибировать функцию NKCC1, вследствие чего обратимо нарушается функция среднего уха. Кроме того, важна роль К+-каналов KCNQ1/KCNE1 (разд. 2): при врожденном дефекте каналов KCNQ1/KCNE1 (синдром Джервелла—Ланге-Нильсена) наблюдается глухота, часто сопряженная с удлиненным интервалом QT на ЭКГ (синдром удлиненного интервала QT 1).
д) Абсорбция и секреция HCO3-. Абсорбция HCO3-, секреция HCO3-, а также абсорбция Na+ и Cl- осуществляются с помощью различных транспортеров.
В транспорте могут участвовать (рис. 4):
- Na+/Н+-антипортеры NHE3 или NHE1;
- фермент карбоангидраза (СА);
- HCO3-/Cl--антипортер АЕ2;
- Na+ HCO3--симпортер NBC1.
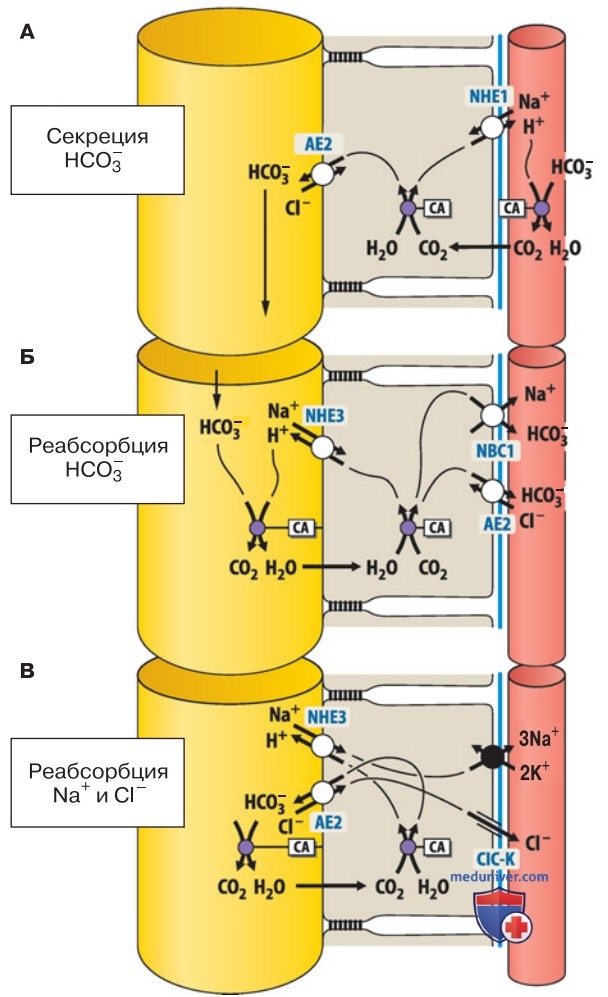
В зависимости от присутствия транспортеров возможны следующие процессы.
1. Секреция HCO3- осуществляется, в частности, в протоках слюнных желез и поджелудочной железы, а также в печени и париетальных (обкладочных) клетках желудка (рис. 4А). Абсорбция HCO3- через базолатеральную мембрану не требует участия HCO3--транспортера, поскольку Н+ выводится в межклеточное пространство с помощью Na+/H+-антипортера (NHE1).
Карбоангидраза катализирует реакцию Н+ + HCO3- с образованием СО2 и Н2О. СО2 диффундирует через плазматическую мембрану в клетку, где благодаря каталитическому действию внутриклеточной СА вновь образуются Н+ и HCO3-. И наконец, третично-активный Cl-/HCO3--антипортер (АЕ2) апикальной мембраны выводит бикарбонат в полость органа.
2. Реабсорбция HCO3-. Реабсорбция HCO3- и секреция Н+ свойственны клеткам проксимальных почечных канальцев и париетальных (обкладочных) клеток желудка (рис. 4Б), причем в желудке секреция Н+ выполняет важную функцию. В эпителиальных клетках, секретирующих HCO3-, два переносчика расположены зеркально по отношению друг к другу.
Н+ выделяется в полость с помощью Na+/H+-антипортера NHE3 апикальной мембраны и при участии СА всасывается обратно в клетку в виде СО2 + Н2О (исключение составляет желудок). Через базолатеральную мембрану HCO3- может транспортироваться посредством двух механизмов.
В некоторых эпителиальных тканях работает Cl-/HCO3--антипортер АЕ2, в других — Nа+,HCO3--симnортер NВС1, причем в последнем случае Na+ выводится из клетки благодаря электрохимическому градиенту HCO3-.
3. Абсорбция Na+ и Cl- (рис. 4). Третий тип расположения выявлен в толстой кишке и желчном пузыре. Здесь в апикальной мембране находятся Na+/Н+-антипортер и Cl-/HCO3--антипортер, благодаря которым Н+ и HCO3- выделяются в полость.
Далее HCO3- под влиянием СА временно преобразуется в СО2 и Н2О; таким образом в клетке снова присутствуют Н+ и HCO3- и вступает в действие антипортер. В то же время Na+ и Cl- входят через апикальную мембрану и могут выводиться через базолатеральный домен.
е) Секреция внутриглазной жидкости. В образовании внутриглазной жидкости участвуют антипортеры NHE1 и АЕ2. При глаукоме следует понижать давление путем ингибирования активности карбоангидразы.
1. Внутриглазная жидкость продуцируется эпителием цилиарного (ресничного) тела. Инициирующая роль принадлежит секреции Na+, Cl-, HCO3- и аминокислот, за которой следует осмос воды. При этом главную функцию выполняют два транспортера — Na+/Н+-антипортер NHE1 и Cl-/HCO3--антипортер АЕ2 (рис. 4А, Б), деятельность которых зависит от активности карбоангидразы.
2. Глаукома («зеленая вода»). При нарушенном балансе между секрецией внутриглазной жидкости и ее оттоком повышается внутриглазное давление; возникает риск повреждения сетчатки и зрительного нерва. В качестве лекарственных средств можно применять ингибиторы карбоангидразы, подавляющие работу NHE1 и АЕ2. В итоге образуется меньше внутриглазной жидкости и давление снижается.
ж) Коротко. Расположение транспортеров в мембране эпителиальных клеток. Эпителий различных органов характеризуется определенной локализацией каналов, переносчиков, насосов и плотных контактов. Na+/К+-АТФаза находится в базолатеральном домене клеточной мембраны.
Переносчики, обеспечивающие поступление в клетку питательных веществ в виде углеводов и аминокислот, в клетках проксимальных («рыхлых») отделов эпителия трубкообразных органов (например, тонкой кишки и проксимальных почечных канальцев) обычно расположены в обоих мембранных доменах, в отличие от размещения в дистальных сегментах этих органов.
Что касается ионных и водных каналов, регулируемых гормонами (альдостероном и вазопрессином) либо внутриклеточными медиаторами (цАМФ или Са2+) и необходимых для поддержания постоянства внутренней среды, то они, наоборот, расположены в апикальных доменах клеточных мембран дистальных («плотных») отделов эпителия (например, в толстой кишке, дистальных почечных канальцах и семявыносящем протоке). Одни и те же транспортеры в зависимости от их локализации могут обеспечивать различные эффекты.
- Вернуться в раздел "Физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 20.8.2024