
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Сигналы клетки, опосредуемые кальцием - с точки зрения физиологии человека
Оглавление статьи:- Повышение концентрации Са2+ в цитозоле в качестве сигнала
- Действие Са2+
- Коротко - кальцийпроводимые сигналы
- Список использованной литературы
а) Повышение концентрации Са2+ в цитозоле в качестве сигнала.
P.S. Кальций (Са2+) высвобождается из внутриклеточных полостей и устремляется в клетку по ионным каналам клеточной мембраны. Эти каналы могут быть потенциалзависимыми или лигандуправляемыми.
1. Высвобождение Са2+. Чтобы повысить концентрацию Са2+ в цитозоле, рецепторы стимулируют фосфолипазу С (ФЛСβ или ФЛСγ). Фосфолипаза отщепляет от определенных фосфолипидов мембраны (фосфатидилинозитолфосфатов) инозитолтрифосфат (IP3) (рис. ниже).
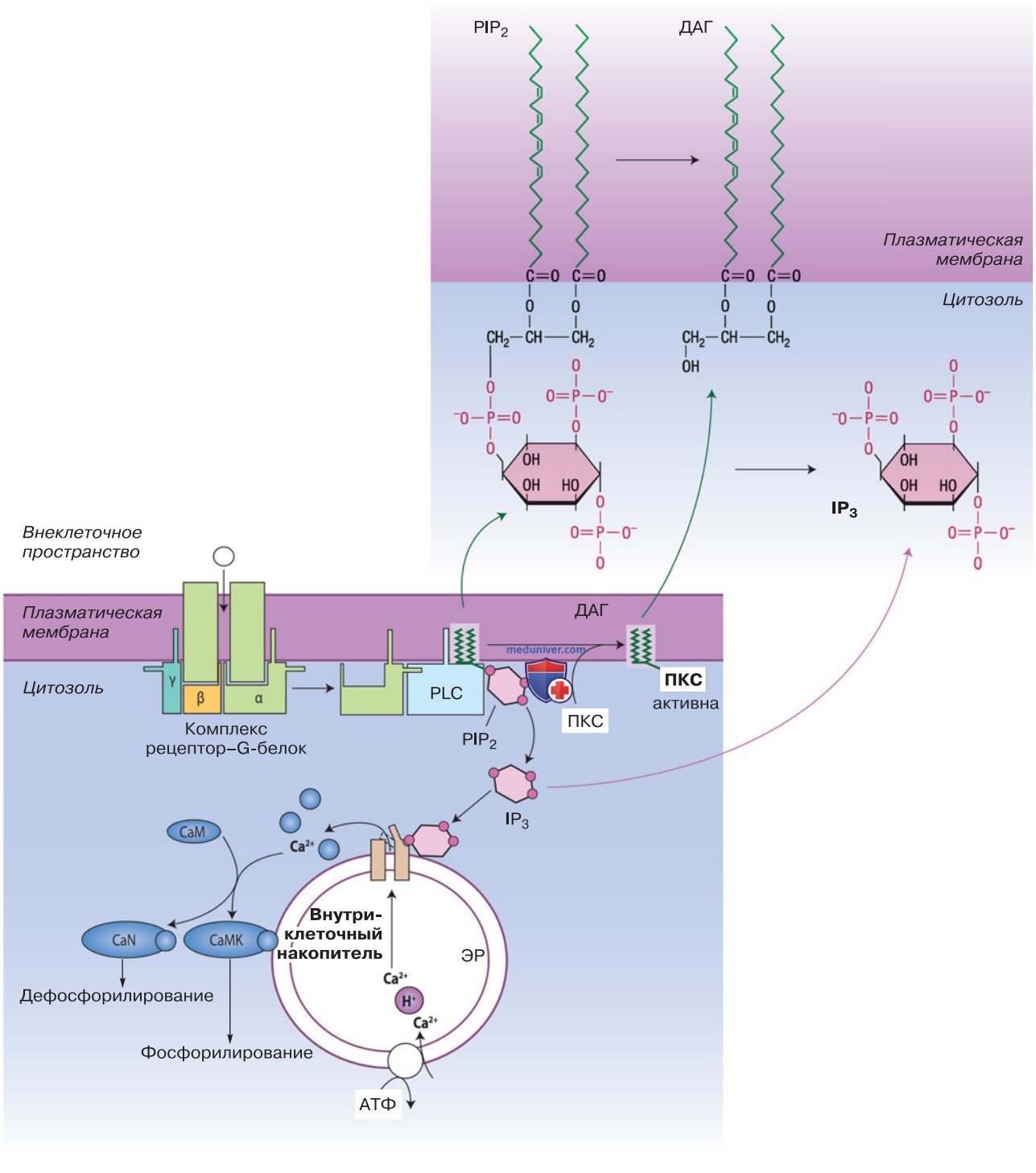
IP3 присоединяется к каналам внутри эндоплазматического ретикулума, которые способствуют высвобождению Са2+ из эндоплазматического ретикулума в цитоплазму. Опорожнение внутриклеточных накопителей Са2+ приводит к активации кальциевых каналов в клеточной мембране, или CRAC (calcium release activated calcium channel), через которые в цитозоль проникает еще большее количество Са2+. CRAC состоит из белка канала Orai и активирующего белка Stim.
2. Диацилглицерол и протеинкиназа С. При отщеплении IP3 из фосфолипидов мембраны образуется диацилглицерол. Вместе с Са2+ диацилглицерол активирует протеинкиназу С (ПКС), которая регулирует транспортные белки, расположенные в клеточной мембране.
Таким образом, ПКС стимулирует обмен Na+/H+ в NHE1 и тем самым снижает концентрацию Н+ в цитозоле. ПКС регулирует дальнейшее разветвление цитоскелета, а кроме того, активируя факторы транскрипции, контролирует синтез белков (рис. выше). Регулируемые с помощью ПКС факторы транскрипции в первую очередь контролируют ранние гены, осуществляющие быстрое приспособление клетки к изменяющимся условиям окружающей среды.
3. Лигандуправляемые и потенциалзависимые кальциевые каналы. Внутриклеточная концентрация Са2+ может первично повышаться за счет притока кальция по ионным каналам, благодаря чему некоторые типы нейромедиаторов могут соединяться с каналами, пропускающими Са2+, и открывать их.
В результате этого так называемые возбудимые клетки располагают потенциалзависимыми проницаемыми кальциевыми каналами, активность которых регулируется посредством разницы потенциала через клеточную мембрану. При нормальной поляризации клеточной мембраны (наибольшее значение составляет -60 мВ) каналы закрыты, при деполяризации они активируются. По каналам передача сигнала между клетками воздействует на потенциал клеточной мембраны.
P.S. Са2+ влияет на активность и экспрессию эффекторных молекул с помощью кальмодулина/ кальцинейрина или путем непосредственного соединения.
1. Кальмодулин и кальцинейрин. Вместе с ПКС Са2+ соединяется с внутриклеточным рецептором кальция кальмодулином (рис. выше). В результате соединения Са2+ и кальмодулина происходит конформационное изменение кальмодулина, которое может стимулировать фосфатазу кальцинейрина. Важнейшим компонентом сигнального пути кальцинейрина является фактор транскрипции НФАТ (нуклеарный фактор активируемых Т-лимфоцитов).
Кальцинейрин дефосфорилирует НФАТ, который в дефосфорилированном состоянии проникает в ядро из цитозоля и стимулирует там транскрипцию генов.
2. Кальцийзависимые функции. Са2+ регулирует множество функций клетки, например сокращение мышц, определяет состояние цитоскелета, осуществляет регуляцию ферментов, принимающих участие в обмене веществ (например, распад гликогена), слияние везикул с мембраной клетки и тем самым выброс нейромедиаторов и гормонов, экспрессию генов, важных для пролиферации клетки, а также активацию ферментов, способных вызвать генетически запрограммированную гибель клетки (апоптоз). Некоторые примеры кальцийзависимой регуляции представлены в табл. 2.

3. Специфика Са2+-сигналов. Из большинства кальцийзависимых функций клетки лишь незначительная часть реализуется в клетке, ведь Са2+ не может стимулировать одновременно и деление клетки и ее смерть. Специфика действия, оказываемого Са2+, определяется исходным состоянием клетки, т. е. одновременно и воздействующими на клетку другими сигналами, и снабжением эффекторными молекулами.
Исходя из этого временная последовательность Са2+-сигналов приобретает большое значение. С одной стороны, кальциевые осцилляции, при которых концентрация внутриклеточного Са2+ краткосрочно и нестабильно повышается (например, каждую минуту на несколько секунд), вызывают такие процессы, как экспрессия генов, необходимая для пролиферации клетки.
С другой стороны, в клеточной мембране происходит разрушение липидных структур, за которым следует перемещение фосфатидилсерина, а также накопление Са2+ в митохондриях с последующей митохондриальной деполяризацией. Это приводит к продолжительному повышению концентрации Са2+ и апоптозу клетки.
в) Коротко - кальцийпроводимые сигналы. Активность фосфолипазы индуцирует образование IP3 и ДАГ. IP3 осуществляет выход Са2+ из внутриклеточных полостей. Опорожнение накопителей активирует кальциевые каналы клеточной мембраны (CRAC). Другие кальциевые каналы в мембране могут активироваться с помощью лигандов или посредством процесса деполяризации.
Повышение концентрации кальция цитозоля выступает в роли сигнала. При этом выполняется целый ряд кальцийзависимых функций клетки:
• В сочетании с другими молекулами Са2+ прямо или косвенно регулирует ПКС, кальмодулин, кальцинейрин и факторы транскрипции.
• Са2+ среди прочих регулирует такие процессы, как сокращение мышц, выброс медиаторов или гормонов, обмен веществ, пролиферацию клетки и апоптоз.
Кальцийзависимые функции определяет временная последовательность сигналов Са2+.
- Рекомендуем ознакомиться далее "Регуляция пролиферации и гибели клетки - с точки зрения физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 20.8.2024