
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Взаимодействие транспортной и барьерной функций эпителиев - с точки зрения физиологии человека
Оглавление статьи:- Структура эпителия
- Межклеточные соединения
- Рыхлость эпителия
- Функциональная организация эпителиев
- Коротко
- Список использованной литературы
P.S. Эпителиальные ткани разграничивают полости организма, содержащие жидкость. Благодаря полярной организации клеток эпителий обеспечивает всасывание и секрецию; оба процесса могут происходить трансклеточно и параклеточно.
1. Функциональный наружный покров. Эпителиальные ткани отделяют организм от окружающей среды, а также образуют внутри него барьеры для полостей, содержащих жидкость. При этом понятие «наружный покров» подразумевает не только кожу тела, но и «функциональные наружные покровы» полостей органов, доступные для поступления веществ из внешней среды.
Некоторые из этих органов получают вещества извне, тогда как другие выводят свое содержимое наружу, например желудочно-кишечный тракт, почечные канальцы, мочевыводящие пути, потовые и слюнные железы.
Внутри организма эпителиальные слои ограничивают и такие содержащие жидкость полости, которые не имеют связи с окружающей средой, например плевра, брюшина (peritoneum), эпикард, перикард и эпителиальная выстилка органов, в частности кровеносных сосудов. В большинстве органов такая выстилка с функциональной точки зрения представляет собой высокопроницаемый эпителий. Выстилка кровеносных сосудов называется эндотелием.
2. Строение эпителия. Клетки эпителия характеризуются полярной организацией и соединены друг с другом специализированными контактами (рис. 1). Апикальный домен клеточной мембраны, как указывает само название, в функциональном смысле обращен к внешней среде. Во многих типах эпителия эта часть поверхности клеточной мембраны образует пальцеобразные выпячивания — микроворсинки, в связи с чем ее называют щеточной каемкой.
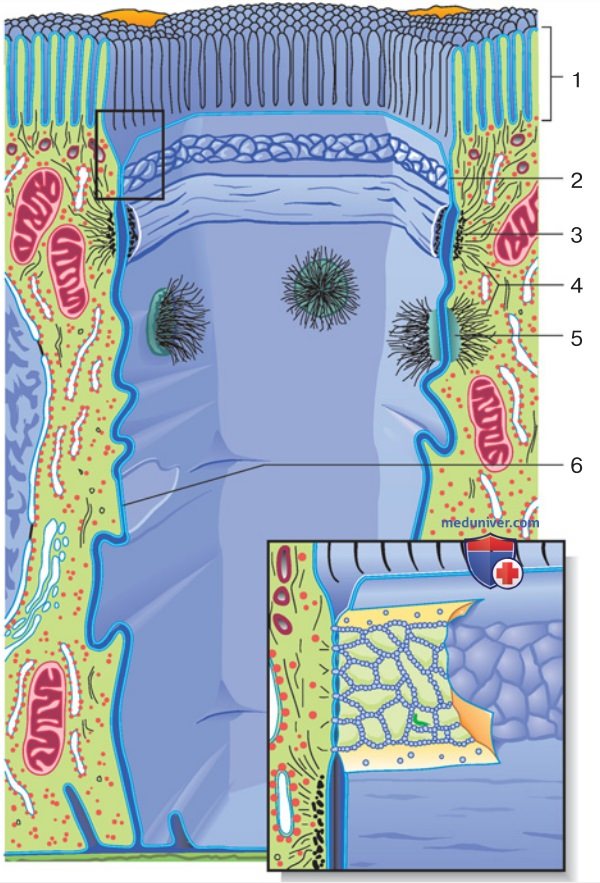
3. Базолатеральный домен плазматической мембраны состоит из базальной части, обращенной к кровеносному сосуду, и латеральных частей. Общее название «базолатеральная плазматическая мембрана» подразумевает, что ее компоненты обладают однотипными транспортерами и не отделены от интерстициального пространства (межклеточного матрикса) каким-либо барьером. Базальная мембрана направляет рост клетки и поддерживает ее целостность, не препятствуя при этом трансэпителиальному транспорту.
4. Полярные свойства. Многие рецепторы и транспортеры ткани после их синтеза в эпителиальной клетке встраиваются в мембранные везикулы ближайшего участка клеточной мембраны, т. е. апикального или базолатерального домена. Так активируемые альдостероном эпителиальные Na+-каналы (ENaC) всегда оказываются в составе апикального домена, а Na+/K+-АТФаза — в составе базолатерального.
5. Пути и направления транспорта. Трансмембранные перемещения веществ в клетку и из нее означают вход и выход соответственно. Транспорт через функциональный внешний покров в межклеточное пространство называется абсорбцией, а в обратном направлении — секрецией. Существует два пути трансэпителиального транспорта.
- Трансклеточный путь: вещества проходят через апикальную и базолатеральную мембрану эпителиальной клетки, а также через участок межклеточной щели.
- Параклеточный путь: вещества проходят через плотный контакт на всем протяжении межклеточной щели.
P.S. Плотные контакты (tight junctions) создают барьер между эпителиальными клетками, но при этом участвуют в параклеточном транспорте.
1. Структура. Между латеральными мембранами смежных клеток имеется межклеточная щель, а также три типа межклеточных соединений (рис. 1): плотные контакты, десмосомы и коннексоны. В то время как десмосомы и коннексоны свойственны и для других типов клеток, плотные контакты (tight junction), или зоны замыкания (zonula occludens), характерны для эпителиев, осуществляющих барьерную функцию.
Плотные контакты расположены ближе к наружной (обращенной к просвету органа) стороне эпителиального пласта и разграничивают апикальную и латеральную части плазматической мембраны.
2. Белки плотных контактов. Сетчатая структура плотных (замыкающих) контактов (рис. 1) образована белками четырех групп: окклюдином, трицеллюлином, белками семейства клаудинов (24 представителя) и молекулами межклеточной адгезии (junctional adhesion molecule, JAM). Эти белки связаны с цитоскелетом через посредство внутриклеточных белков (в том числе ZO-1, ZO-2 и ZO-3).
За исключением JAM, белки плотных контактов состоят из четырех трансмембранных доменов, а также одной внутриклеточной и двух внеклеточных петель ECL1 и ECL2. Благодаря заряду своих аминокислотных остатков ECL1 определяет барьерные свойства (т. е. уровень проницаемости) контакта, тогда как ECL2 выполняет опорную и, в некоторых случаях, рецепторную функцию.
2. Функциональные свойства. Плотные контакты осуществляют два вида барьерных функций:
- препятствуют латеральной диффузии мембранных белков, в частности перемещению апикальных белков к базолатеральной мембране и наоборот;
- создают барьер для трансэпителиального транспорта. В некоторых эпителиях этот барьер практически непроницаем, однако в других случаях межклеточный контакт может пропускать ионы даже легче, чем плазматическая мембрана. Проницаемость плотных контактов определяется, с одной стороны, морфологическими факторами, т. е. шириной «ячеек» их сети (рис. 1, вставка), а с другой стороны, молекулярными свойствами взаимодействующих белков.
Большинство белков плотного контакта ассоциированы с его барьерной функцией. Сюда относятся клаудин-1, -4, -5, -8, -14, -19, а также окклюдин. Особая барьерная роль принадлежит трицеллюлину, который затрудняет проникновение макромолекул в участках контакта между тремя эпителиальными клетками.
Некоторые клаудины выполняют противоположную функцию: вместе с клаудинами смежной клетки они составляют параклеточные каналы. Наиболее явно это свойство выражено у белка клаудина-2, который образует параклеточный канал для малых катионов и присутствует преимущественно в эпителиях с проницаемыми плотными контактами (см. табл. А1). Клаудин-10b можно обнаружить, как правило, в дистальных почечных канальцах, где он формирует катионные каналы, тогда как сплайс-вариант клаудин-10а характерен главным образом для анионных каналов в проксимальных почечных канальцах.
Клаудин-16, локализованный в восходящей части петли Генле и начальном участке дистального почечного канальца, пропускает катионы и опосредованным образом стимулирует трансклеточную абсорбцию Mg2+ за счет индукции бестрофина — трансмембранного белка хлорного канала.
3. Нарушения эпителиальных барьеров. Эти нарушения выявляются при многих заболеваниях и даже могут лежать в основе патологического процесса. К подобным патологиям относятся воспаления кишечника, например язвенный колит и болезнь Крона; энтеропатогенные инфекции (например, нарушение структуры плотных контактов под действием холерного токсина); аутоиммунная реакция на некоторые продукты питания (например непереносимость глютена при целиакии); гиперчувствительность к некоторым лекарственным препаратам (в частности, к нестероидным противовоспалительным средствам, или НПВС). Иарушения эпителиальных барьеров приводят к:
- патологически повышенному выведению растворенных низкомолекулярных веществ и воды;
- всасыванию вредного фактора с последующим его поступлением в общий кровоток; затем вредный фактор продолжает через кровь оказывать свое неблагоприятное действие в том числе и на эпителиальные ткани, т. е. возникает «порочный круг».
P.S. Болезнь Крона. Болезнь Крона — хроническое воспалительное заболевание, поражающее эпителиальный слой стенок кишечника и способное распространяться на другие, не связанные между собой участки пищеварительного тракта. Вероятная причина заболевания — продолжительная активация системы интестинальной иммунной защиты у генетически предрасположенных к этому людей.
К симптомам и признакам относятся в первую очередь диарея, абдоминальные боли, лихорадка, снижение массы тела, появление фистул в области заднего прохода. Большинство симптомов заболевания обусловлено образованием провоспалительных цитокинов, таких как фактор некроза опухоли-α (tumornecreose-factor, TNFα) и γ-интерферон.
- Патофизиологические механизмы. Важное значение имеет нарушение барьерной функции эпителия кишечника в результате локального язвенного процесса, интенсивного апоптоза и повреждения плотных контактов. TNFα уменьшает содержание уплотняющего контакты белка клаудина-8 и увеличивает содержание клаудина-2, проницаемого для катионов. Нарушение барьера имеет два последствия: во-первых, из просвета кишечника интенсивно всасываются вредные вещества, усугубляющие патологический процесс, во-вторых, в просвет выделяется большое количество растворенных веществ и воды, что приводит к диарее. При лечении необходимо использовать иммунодепрессанты.
P.S. Десмосомы. Опоясывающая десмосома (англ. аdherens junction, лат. zonula adheаrens) обеспечивает механическое сцепление эпителиальных клеток и вместе с плотным контактом составляет соединительный комплекс (англ. junctional complex).
4. Коннексон, третий тип межклеточных соединений, образован коннексинами — каналами, пронизывающими плазматические мембраны двух смежных клеток. Коннексоны входят в состав щелевых контактов (нексусов). Значительное повышение внутриклеточной концентрации Са2+, например в результате разрушения плазматической мембраны, вызывает закрывание коннексонов и отгораживание соседних интактных клеток от пострадавших клеток. Благодаря этому зона повреждения не распространяется, например при инфаркте миокарда.
В апикальной области межклеточная щель более или менее перекрыта соединительными комплексами, тогда как в базальной области подобные ограничивающие структуры отсутствуют. При невысокой скорости транспорта или секреции межклеточная щель остается узкой, однако в случае усиленной абсорбции может существенно расшириться. Что касается нарушений диффузии, даже незначительных, то оно может произойти лишь при крайней степени сужения щели.
P.S. Отношение параклеточной проницаемости к трансклеточной проницаемости служит показателем «рыхлости» эпителия.
Плотный контакт, вопреки своему названию, до некоторой степени проницаем, когда речь идет о трансэпителиальном транспорте; по существу, это соответствует параклеточной пропускной способности. Что касается трансклеточной проницаемости, то она зависит от проницаемости обоих доменов плазматической мембраны. Индекс проницаемости плотного контакта и плазматической мембраны служит показателем рыхлости эпителия. По индексу проницаемости различают три уровня (класса) рыхлости эпителия.
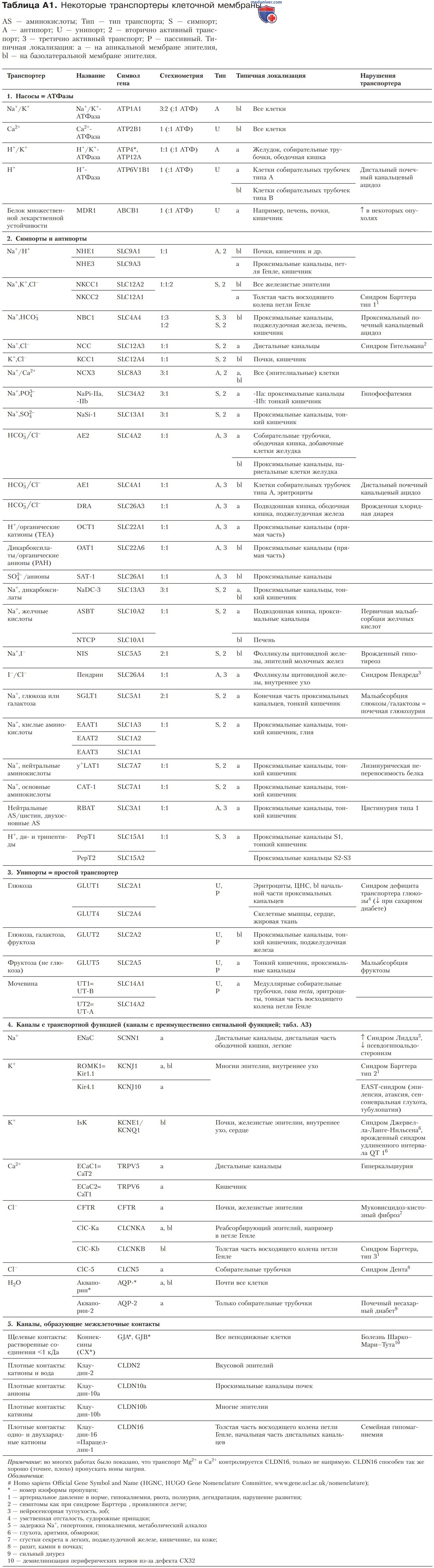
Основу классификации эпителиев (табл. 2) составляют отношения электрической проводимости, а также проницаемости к ионам Na+, K+ и Cl-. В случае более крупных молекул электролитов или воды показатели рыхлости могут отличаться от значений, приведенных в таблице.
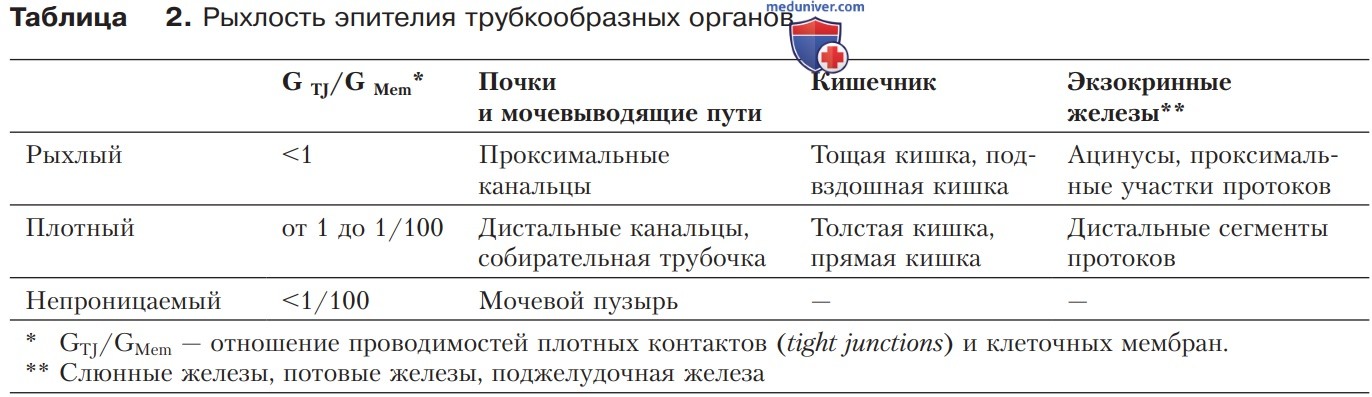
1. Непроницаемые эпителии. Они практически лишены транспортной функции и прежде всего выполняют роль барьеров. Сюда относятся только эпителий мочевого пузыря и эпидермис.
2. Плотные эпителии. Что касается количественных характеристик транспорта веществ, то плотные эпителии, как правило, обладают слабой пропускной способностью, однако могут создавать значительные градиенты. Подобными свойствами отличаются дистальные сегменты эпителиев всех трубкообразных органов, например дистальных почечных канальцев, семявыносящих протоков, толстой и прямой кишки, дистальных сегментов выводных протоков поджелудочной железы, слюнных и потовых желез. По определению плотные контакты эпителиев этого типа менее проницаемы, чем клеточные мембраны.
Следовательно, здесь трансэпителиальный транспорт является преимущественно трансклеточным и лишь отчасти параклеточным. В таких эпителиях скорость транспорта регулируется в широких пределах, в частности посредством гормонов. В итоге транспорт может осуществляться против градиента, даже если он высокий.
3. Гематоэнцефалический барьер. Эндотелий капилляров головного мозга существенно плотнее, чем у большинства других капилляров, и по свойствам близок к плотному эпителию. Стенки капилляров головного мозга лишены отверстий, а плотные контакты обладают здесь меньшей пропускной способностью, чем клеточные мембраны. Поэтому полярные молекулы, лишенные транспортеров, вообще не способны выходить в мозг либо поступают в небольших количествах. В то же время полярные молекулы, обладающие мембранными транспортерами, могут перемещаться даже против электрохимических градиентов.
4. Рыхлые эпителии. Характеризуются интенсивным транспортом, причем мелкие молекулы растворенных веществ проникают независимо от их концентрации. Плотные контакты здесь обладают большей пропускной способностью, чем клеточные мембраны. К рыхлым эпителиям относятся проксимальные сегменты эпителиев всех трубкообразных органов, например проксимальных почечных канальцев, тонкой кишки, желчного пузыря, ацинусов и проксимальных сегментов выводного протока поджелудочной железы, а также выводных протоков слюнных и потовых желез.
У эпителиев этого типа особенно высокие уровни абсолютной проницаемости и трансклеточного транспорта. В результате осмоса через мембрану поступает вода вместе с частицами растворенных веществ (перенос растворенного вещества вместе с растворителем — solvent drag). В итоге благодаря осмотическому градиенту транспортируется некоторое количество частиц без дополнительных затрат метаболической энергии.
г) Функциональная организация эпителиев.
P.S. Выведение конечных продуктов метаболизма через эпителий трубкообразных органов подчиняется единой схеме: в проксимальных отделах относительно большие количества веществ транспортируются против низких градиентов, а в дистальных — малые количества веществ против высоких градиентов.
1. Сегментарная гетерогенность. Плотность эпителия почек, кишечника и выводных протоков экзокринных желез возрастает по мере перехода от проксимальных к дистальным сегментам (табл. 2). Подобная сегментарная гетерогенность соответствует единой схеме формирования выводимого продукта во всех трех рассматриваемых типах эпителия — рыхлом, сравнительно плотном и практически непроницаемом.
- Выработка изоосмотического первичного фильтрата. Первичный продукт, поступающий в просвет органа, изоосмотичен плазме крови (при ультрафильтрации в почечных клубочках и первичной секреции в ацинусах экзокринных желез) и/или вырабатывается путем осмоса в результате потока воды (в желудке и в начальном отделе эпителия всех трубкообразных органов).
- Изоосмотический транспорт. Через рыхлый эпителий проксимальных сегментов изоосмотически транспортируются большие количества растворенных веществ и воды без существенного участия гормонов.
- Обработка выводимого продукта. Через относительно плотный эпителий дистальных сегментов транспортируются лишь небольшие количества веществ, но против высоких электрохимических градиентов. Транспорт эффективно регулируют гормоны. В результате выводимый продукт вырабатывается с определенной концентрацией и массой веществ, что соответствует поддержанию постоянства внутренней среды организма. При снижении рыхлости эпителия дистальных сегментов возрастает способность к переносу веществ против градиентов.
- Накопление выводимого продукта. Эпителий мочевого пузыря практически не осуществляет транспорт, но способен длительно поддерживать очень высокие градиенты между просветом органа и кровью. Таким образом, мочевой пузырь выполняет накопительную функцию.
д) Коротко. Транспортные и барьерные функции. Эпителиальные клетки соединены между собой плотными контактами, создающими барьер для свободного проникновения веществ и воды через мембрану. Апикальный и базолатеральный домены плазматической мембраны обладают своим набором ионных каналов, переносчиков и АТФаз. Границей между двумя доменами клеточной поверхности служат плотные контакты.
Благодаря сочетанию двух функций — транспортной и барьерной — в организме сосуществуют сложно организованные компартменты. Трансэпителиальный транспорт может осуществляться трансклеточным путем — через клеточные мембраны и параклеточным — через плотные контакты. Эти клеточные соединения образованы такими белками, как окклюдин, трицеллюлин и представители семейства клаудинов.
Большинство белков, составляющих плотные контакты, выполняют барьерную роль, однако некоторые из них (в частности, клаудин-2, -10а и -10b) образуют параклеточные поры. При многих заболеваниях нарушения барьерной функции усугубляют патологический процесс.
Проницаемость плотных контактов в эпителиях трубкообразных органов, например кишечника и почечных канальцев, снижается по направлению к дистальному отделу. Таким образом, формирование продукта, выводимого из организма, реализуется по определенной схеме и включает в себя два основных этапа:
• в проксимальных сегментах — нерегулируемый транс- и параклеточный транспорт большой массы вещества против низких градиентов;
• в дистальных сегментах — частично регулируемый гормонами трансклеточный транспорт ограниченной массы вещества против высоких градиентов.
- Рекомендуем ознакомиться далее "Активный и пассивный транспорт клетки - с точки зрения физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 20.8.2024