
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Активный и пассивный транспорт клетки - с точки зрения физиологии человека
Оглавление статьи:- Пассивный транспорт
- Диффузия воды
- Активный транспорт: первичный, вторичный и третичный
- Коротко
- Список использованной литературы
а) Пассивный транспорт. Пассивный транспорт осуществляется за счет градиентов гидростатического давления, концентрации и электрического потенциала.
1. Градиент. Это понятие, которое далее будет часто встречаться, означает уменьшение свободной энергии вещества на протяжении некоторого участка пространства (-dE/dx). Когда речь идет о физиологии транспортных процессов, следует также указывать направление градиента, поскольку в биологических системах перемещение может быть как «нисходящим» («сверху вниз», или «с горы») — по градиенту (пассивный транспорт), так и «восходящим» («снизу вверх», или «в гору») — против градиента (активный транспорт).
На схемах транспорта можно видеть стрелки (табл. 1), указывающие направление и угол наклона электрохимических градиентов.

2. Механизмы активного и пассивного транспорта. Обе формы транспорта требуют затрат энергии, которая обеспечивается за счет либо гидролиза АТФ, либо физических градиентов.
- Активный транспорт может идти против внешних градиентов, т. е. «в гору».
- Пассивный транспорт всегда идет по внешним градиентам, т. е. «с горы».
Разделение транспорта на активный и пассивный позволяет прежде всего выделить перемещения, опосредуемые транспортными белками. В отличие от них, диффузия через липидную фазу клеточной мембраны, а также параклеточный транспорт через плотные контакты и межклеточную щель являются пассивными.
3. Фильтрация и ультрафильтрация. Транспорт, обусловленный градиентом гидростатического давления, происходит через фильтры. Уровень фильтрации линейно зависит от движущей силы. Поры фильтров, используемых в быту (например, кофейного фильтра), отделяют нерастворенные частицы от растворенных.
Однако поры фенестрированного эндотелия капилляров еще меньше по диаметру и не пропускают крупные молекулы даже растворенных веществ. Такой процесс в эндотелии капилляров, называемый ультрафильтрацией, представляет собой важнейший транспортный механизм, проницаемость которого очень высока. Однако ультрафильтрация практически нулевая в более плотных клеточных мембранах, а также в эпителиальных тканях, соответствующих более узкому пониманию этого термина.
Относительно ультрафильтрации (а также применительно к переносу растворенного вещества вместе с растворителем — solvent drag) существует предположение, что увлекаемые водой частицы могут либо пройти через водопроницаемые участки, либо «отсеяться» (отфильтроваться). Этот процесс количественно характеризуется коэффициентом фильтрации S (табл. 3), значения которого могут варьироваться от 0 (отсутствие проникновения частиц) до 1 (беспрепятственное проникновение). Формально за величину S принимают вероятность прохождения частиц через мембрану.

4. Диффузия. Перенос частиц из области высокой концентрации туда, где концентрация ниже, называют диффузией. Направление переноса объясняется следующим образом. В процессе хаотичного броуновского движения частицы сталкиваются особенно часто в зоне высокой концентрации, так что они уступают друг другу место, перемещаясь в область более низкой концентрации. Таким образом, движущей силой диффузии служит градиент концентрации. Диффузия незаряженных (неионизированных) частиц — неионная диффузия описывается уравнением (1) в табл. 3.
5. Электрохимический градиент. Диффузия заряженных частиц определяется градиентом не только концентрации, но и электрического потенциала (табл. 3, уравн. (2). Эти два вида градиентов в совокупности составляют электрохимический градиент. Взаимосвязь между ионной проницаемостью Р, ионной проводимостью G и специфическим сопротивлением мембраны R представлена уравнением (3) в табл. 3.
6. Уравнение Нернста. Уравнение характеризует соотношение между электродвижущей силой и ионными концентрациями некоторых ионов (табл. 3, уравн. (4). Рассчитав постоянную Фарадея, получаем упрощенное уравнение (4а). Из него следует, что если, например, трансмембранное соотношение концентраций одновалентного иона соответствует 10:1, то мембранный потенциал равен 61 мВ. Если концентрации иона по обе стороны мембраны одинаковы, транспорт иона равен нулю. При этом ионы этого вида распределены пассивно в соответствии с электрохимическим градиентом, а мембранный потенциал равен равновесному потенциалу для соответствующего иона.
7. Простая диффузия. Такая диффузия не требует участия транспортных белков. Частицы проникают через двойной фосфолипидный слой мембраны либо перемещаются в свободной жидкости, причем это происходит без насыщения. В случае простой диффузии через липидную фазу клеточной мембраны проницаемость транспортируемых молекул пропорциональна их липофильности. Через липидную фазу клеточной мембраны диффундируют прежде всего газы (например О2, СО2, N2), слабые электролиты в незаряженной форме и другие неполярные вещества, тогда как вода и ионы диффундируют очень слабо либо не диффундируют совсем.
P.S. Облегченная диффузия. Это понятие возникло раньше, чем были открыты транспортные белки, и объединяет все формы диффузии, опосредуемые такими белками. Облегченная диффузия происходит с насыщением. Иногда термин ошибочно применяют в текстах об унипортерах.
б) Диффузия воды. Осмос создает осмотическое давление, из-за которого происходит перенос растворенного вещества вместе с растворителем (растворное привлечение, solvent drag); белки обеспечивают коллоидно-осмотическое давление и эффект Доннана.
1. Осмос. Осмос — это диффузия растворителя (воды). Стимулом служит градиент концентрации, в данном случае — самой воды. Представление о «концентрации воды» достаточно необычно: максимальная концентрация свойственна чистой воде; чем больше веществ в ней растворено, тем значительнее они «вытесняют» воду. Соответственно концентрация воды обратно пропорциональна ее осмотичности.
2. Осмотическое давление. При наличии мембраны, проницаемой для воды и плохо пропускающей частицы растворенного вещества, возникает процесс осмоса. Величина осмотического давления зависит от осмотичности растворителя и проницаемости мембраны к частицам, а направление противоположно устремлению воды.
3. Коллоидно-осмотическое давление. Это доля общего осмотического давления, которую обеспечивают макромолекулы (коллоиды). Онкотическим давлением называется сумма коллоидно-осмотического давления и небольшого дополнительного осмотического давления, возникающего в результате распределения Доннана (см. далее).
Благодаря локальному осмосу вода пассивно следует вместе с растворенными в ней веществами. Как правило, водопроницаемость достаточно высока и вода по существу перемещается изоосмотически (11 на 290 мосмоль/кг).
5. Растворное привлечение (solvent drag). Это явление означает, что поток воды увлекает через мембрану мелкие растворенные частицы. Такое явление характерно, например, для тонкой кишки и проксимальных почечных канальцев.
6. Эффект Доннана. При физиологическом уровне рН анионы представлены в плазме крови только белками. Поскольку в результате ультрафильтрации белковые молекулы задерживаются, все виды ионов, участвующие в этом процессе, распределяются по обе стороны мембраны неравномерно (рис. 1). Подобная ситуация свойственна для всех клеточных мембран, так как цитоплазма богата отрицательно заряженными белками, которые клетка не может вывести.
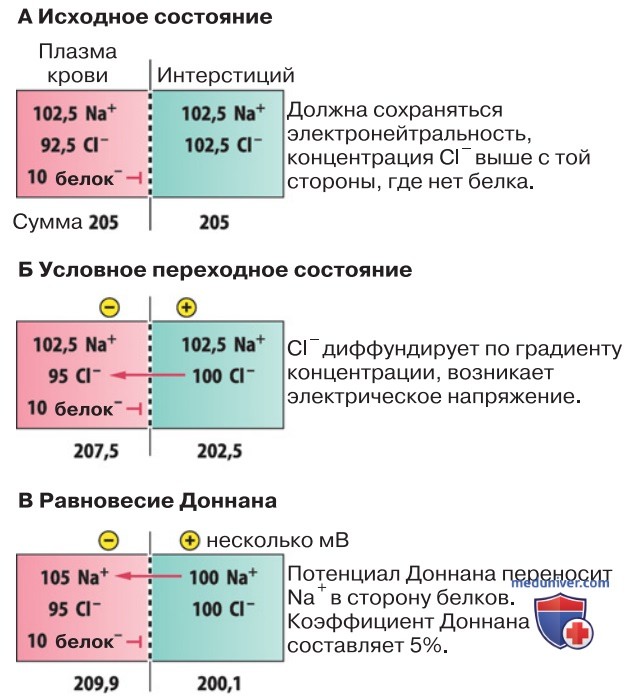
P.S. Распределение Доннана. В результате первичного неравномерного распределения проникающих через мембрану ионов генерируется небольшая трансмембранная разность потенциалов (потенциал Доннана). Мембранный потенциал, в свою очередь, сказывается на конечном распределении Доннана. Чтобы описать распределение Доннана, необходимо использовать фактор Доннана, одинаковый для всех пассивно распределяющихся катионов и анионов. В условиях равновесного распределения концентрация одновалентных катионов в плазме крови на 5% выше, а одновалентных анионов — на 5% ниже, чем в межклеточной жидкости. Для двухвалентных ионов разность концентраций составляет 10%.
в) Активный транспорт: первичный, вторичный и третичный. Первично-активный транспорт требует непосредственных затрат АТФ. Вторично-активный транспорт работает по принципу симпорта или антипорта; обычный стимул — это градиент концентрации Na+. Третично-активный транспорт запускается вторично-активным транспортом.
1. Первично-активный транспорт. Первично-активными транспортерами являются уже упоминавшиеся АТФазы (насосы), способные «перекачивать» растворенные вещества в направлении, противоположном их электрохимическому градиенту, затрачивая при этом метаболическую энергию. Типичный пример первично-активного транспорта представлен на рис. 2А.
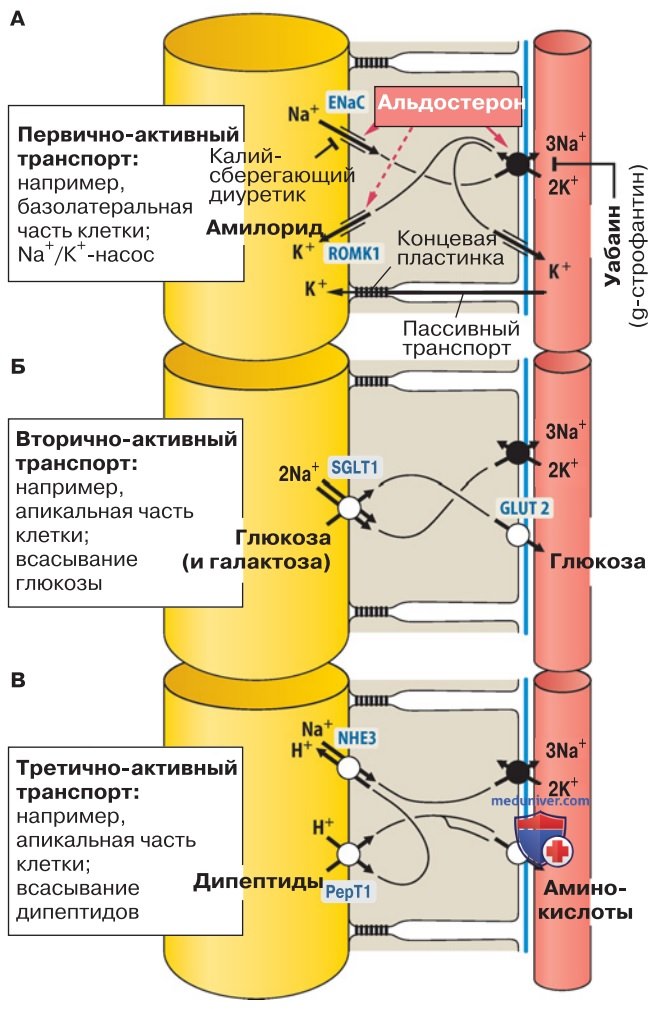
2. Вторично-активный транспорт. Механизм вторично-активного транспорта можно пояснить на примере симпортера SGLT1 — белка-переносчика Na+/глюкозы в апикальном домене клеточных мембран многих типов эпителия (рис. 2Б). SGLT1 осуществляет транспорт только в случае присоединения двух ионов Na+ и одной молекулы глюкозы. При этом частицы одного и другого типа не обязательно должны иметь «нисходящий» градиент; сопряженный транспорт работает по отношению к частицам обоего типа, если сумма их градиентов обеспечивает соответствующее направление.
Поскольку градиент Na+ направлен из внешней среды внутрь клетки, молекулы глюкозы могут всасываться в клетку и против своего градиента концентрации.
P.S. Два необходимых компонента. Симпортер SGLT1 пассивно работает сам по себе, если учесть, что энергия для транспорта глюкозы поступает благодаря электрохимическому градиенту Na+. Следовательно, градиент Na+ должен стабильно поддерживаться с помощью первично-активного транспорта, т. е. работы Na+/K+-АТФазы, содержащейся в базолатеральной мембране. В итоге транспорт глюкозы опосредованно обеспечивается энергетическим метаболизмом клетки.
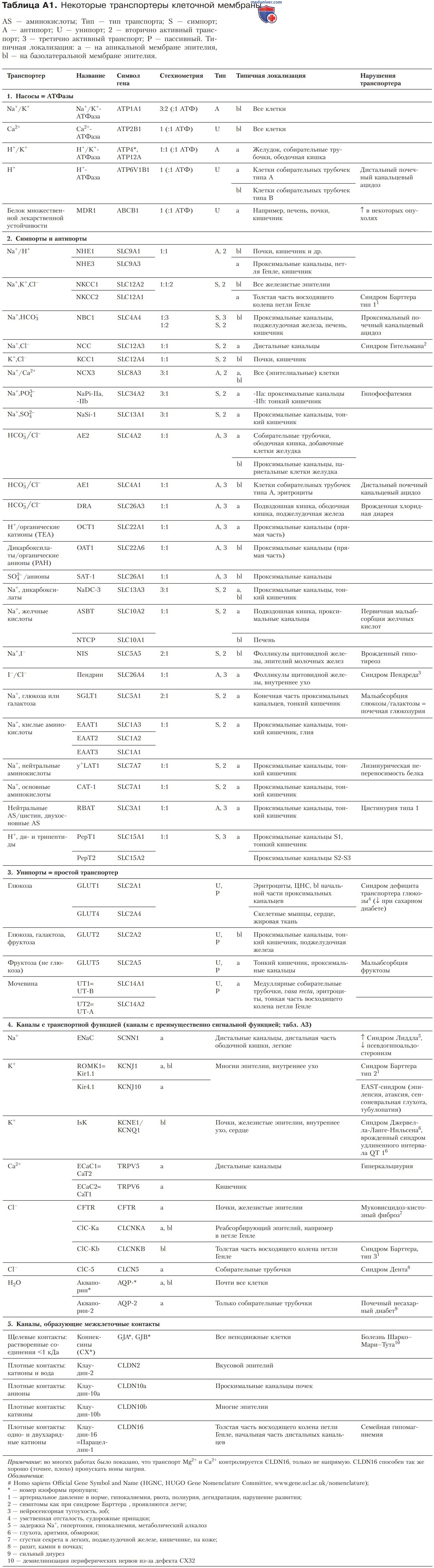
Вторично-активный транспорт распространен достаточно широко; в сопряженном транспорте чаще всего участвует Na+. Наиболее значимые симпортеры и антипортеры представлены в табл. А1.
3. Третично-активный транспорт. Аналогично той ситуации, когда вторично-активный транспорт стимулируется первичным, инициатором третично-активного транспорта является вторичный, например симпортеры Н+/дипептидов (РерТ1 и РерТ2) в апикальной мембране эпителиальных клеток тонкой кишки и проксимальных канальцев (рис. 2В).
Акцептируя дипептиды и трипептиды, эти транспортеры могут переносить их в клетку против электрохимического градиента, если градиент только что присоединенного Н+ направлен в клетку. Такой градиент поддерживается вторично-активным Na+/Н+-антипортером, который содержится в апикальной мембране и, в свою очередь, опосредованно стимулируется Na+/К+-АТФазой.
Третично-активный транспорт тоже встречается довольно часто (см. табл. А1). Особенно разнообразны функции дикарбоксилат/ РАН-антипортера (ОАТ1), который акцептирует многочисленные органические анионы, и Н+/ТЕА-антипортера (ОСТ1), транспортирующего органические катионы.
г) Коротко. Пассивный и активный транспорт. Градиенты отражают разницу концентрации, электрического потенциала или давления между двумя точками пространства.
Процессы транспорта, обусловленные градиентами, называются пассивными и протекают «сверху вниз», т. е. по направлению к области более низкой концентрации транспортируемого вещества.
Сумма градиентов концентрации и электрического потенциала называется электрохимическим градиентом и служит движущей силой диффузии.
Градиент гидростатического давления составляет движущую силу ультрафильтрации и действует только в эндотелии в связи с его высокой водопроницаемостью.
И наоборот, транспорт, осуществляемый «снизу вверх», называется активным или первично-активным. При активном транспорте ионные насосы, например Na+/К+-АТФаза, непосредственно используют для перемещения ионов Na+ и K+ метаболическую энергию гидролиза АТФ. Однако активный транспорт может быть также связан с АТФ опосредованно. В таком случае его называют вторично- или третично-активным транспортом, иначе говоря, сопряженным транспортом (например, Na+ и другого вещества) с участием симпортера или антипортера.
Стимулом сопряженного транспорта служит внутриклеточная концентрация ионов Na+, сниженная в результате первично-активного транспорта. Аналогичным образом вторично-активный транспорт становится пусковым механизмом для третично-активного транспорта.
- Рекомендуем ознакомиться далее "Расположение транспортеров в эпителиальных клетках - с точки зрения физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 20.8.2024