
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Роговой конверт эпидермиса - состав, функции
Наряду со значительным накоплением промежуточных филаментов внутри кератиноцитов, две другие особенности позволяют эпидермису создать крайне эффективный упругий барьер: (1) роговой конверт (образующийся под плазмалеммой белковый полимер с ковалентными перекрестными связями) и (2) внеклеточное гидрофобное вещество, состоящее из специализированных липидов, которые синтезируются терминально дифференцирующимися кератиноцитами. Подробное обсуждение липидов эпидермиса приведено в главе 47 и здесь не дублируется.
Роговой конверт представляет собой сложную структуру, состоящую из соединенных ковалентными перекрестными связями белков, которая образуется под плазмолеммой кератиноцитов в зернистом слое эпидермиса и со временем замещает саму плазмолемму. Данный процесс является составной частью заключительного этапа на пути к полной терминальной дифференцировке. Эта толстая оболочка (20 нм), степень нерастворимости и прочность которой столь высоки, что она способна выдерживать кипение в присутствии сильных денатуратов, покрывает собой внутреннее содержимое клетки и существенно влияет на физико-химические свойства рогового слоя.
Исключительная стабильность рогового конверта во многом обусловлена большим количеством ε-γ-глютамил-лизиновых изопептидных связей между его первичными элементами, усиленных дисульфидными мостиками. Подобные изо-пептидные связи катализируются трансглютаминазами — семейством кальций-зависимых ферментов. Наружная поверхность рогового конверта ковалентно связана с церамидами и другими специализированными липидами, которые производятся и секретируются дифференцированными кератиноцитами зернистого слоя.
Внутренняя поверхность рогового конверта перекрестно связана с крупными пучками плотно упакованных кератиновых промежуточных филаментов.
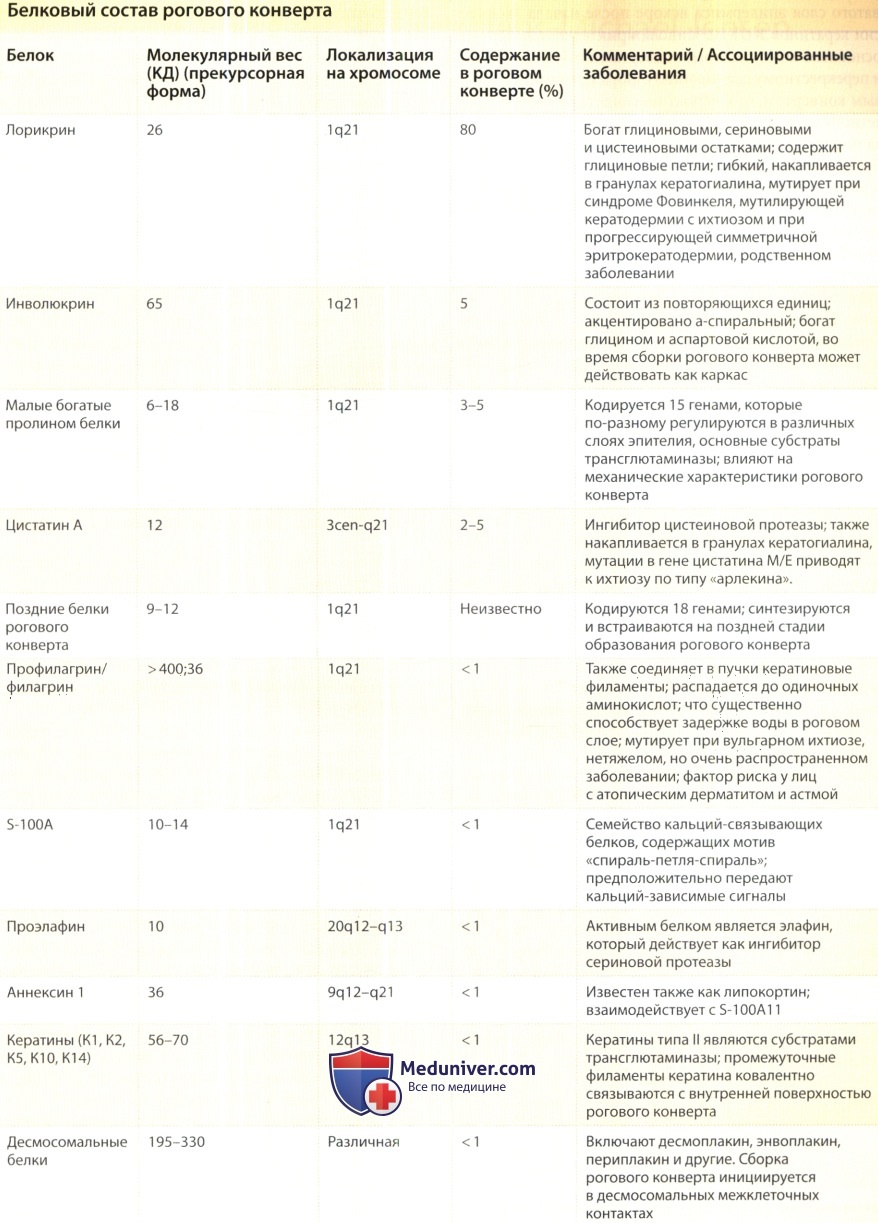
Основными белковыми составляющими рогового конверта являются лорикрин, инволюкрин, филагрин, элафин, цистатин А, корнифелин, несколько малых богатых пролинами белков, кальций-связывающие белки S-100 и «поздние белки рогового конверта». Кроме перечисленных белков, с роговым конвертом перекрестно связаны также ключевые компоненты десмосом (например, десмоплакин, энвоплакин и периплакин), а также несколько кератинов типа II (К1, К2, К5).
Белки рогового конверта обладают рядом любопытных свойств. Во-первых, многие из этих белков кодируются генами, сгруппированными у человека на хромосоме 1q21 в составе локуса, отвечающего за эпидермальную дифференцировку. Играя очевидную эволюционную роль, данное геномное скопление также обусловливает возможность скоординированного регулирования посредством ds-действующих детерминантов.
Во-вторых, ряд таких белков являются прекурсорами, которые во время образования рогового конверта активируются протеолитическим расщеплением, связыванием кальция или другими модификациями. В-третьих, многие белки рогового конверта синтезированы из повторяющихся элементов, у которых отсутствует четкая трехмерная структура. В-четвертых, практически все белки рогового конверта являются субстратами трансглютаминазы.
Хорошо изученными компонентами рогового конверта являются инволюкрин, лорикрин и филагрин. Запуск биосинтеза инволюкрина происходит в дифференцирующихся эпидермальных кератиноцитах шиповатого слоя эпидермиса вскоре после начала экспрессии кератинов К1/К10. Инволюкрин является первым основным компонентом, подвергающимся активации и перекрестному связыванию с формирующимся роговым конвертом, что, вероятно, свидетельствует о «скелетной» роли данного белка, который концентрируется на внешней поверхности зрелого рогового конверта.
Профилагрин и лорикрин синтезируются в виде прекурсоров и накапливаются в гранулах кератогиалина. Лорикрин играет роль главного структурного компонента рогового конверта, составляя около 80% от его веса. Этот белок является в значительной мере неструктурированным, обладая при этом высокой эластичностью, и имеет глициновые петли, характерные для кератинов типа II. Филагрин, активируемый путем дефосфорилирования и протеолитического расщепления, в более высоком зернистом слое эпидермиса участвует в агрегации промежуточных филаментов кератина в пучки (чем объясняется его включение в роговой конверт), а также распадается на свободные аминокислоты, благодаря которым клетки приобретают способность удерживать воду.
Основные сведения о других компонентах рогового конверта приводятся в таблице ниже.
Белковый состав и тонкая структура эпидермального рогового конверта несколько различаются в зависимости от вида многослойного плоского эпителия; значение такой гетерогенности рогового конверта в настоящий момент остается не проясненным.
О том, насколько важна роль рогового конверта в осуществлении барьерной функции эпидермиса, свидетельствуют симптомы, связанные с мутациями первичных составляющих рогового конверта. Это стало особенно очевидно благодаря недавно выясненной роли мутаций филаггрина при вульгарном ихтиозе и атопическом дерматите (АД), а также АД-ассоциированной сенной лихорадке или астме (так называемый «атопический марш»). Частичная или полная утрата белка филаггрина, вызванная мутациями со сдвигом рамки в соответствующем гене, нарушает барьерную функцию эпидермиса и облегчает проникновение в кожу аллергенов и раздражителей, таким образом провоцируя местное (и даже системное) воспаление, сопровождающееся зудом, эритемой и шелушением, являющимся следствием патологии процесса ороговения.
Таким образом, способность эпидермиса обеспечивать барьерную функцию определяет степень влияния на организм экзогенных факторов и интенсивность взаимодействия с окружающей средой.
В последних статьях на сайте рассмотрена концепция стволовых клеток, играющих важнейшие роли в эпидермисе, волосяных фолликулах и других самообновляющихся тканях. Известно, что определенные кератины типов I и II попарно экспрессированы в зависимости от стадий нормальной и патологической дифференцировки эпидермиса, и что мутации основных эпидермальных кератинов являются причиной различных форм пузырных заболеваний кожи. Однако функциональное значение разнообразных кератинов, как в аспекте их структурной роли, так и в плане их участия в сигнальной трансдукции и транспорте органелл (что явилось недавним открытием), находится лишь на начальном этапе изучения.
Дальнейшего исследования требует также роль стволовых и ТА-клеток в пролиферации и гомеостазе эпидермиса, а также понимание, как функционируют филаменты кератина на атомном уровне в нормальном эпидермисе и в патологических условиях. Будущие исследования в данной области должны сформировать новые направления терапии (в частности, такие как, клеточная и генная терапии) ряда тяжелых кожных заболеваний.
- Рекомендуем далее ознакомиться со статьей "Защитная и барьерная функция кожи как органа"
Оглавление темы "Гистология, физиология кожи.":- Клеточная и генная терапия стволовыми клетками эпидермиса
- Классификация кератинов человека
- Кератиновые белки промежуточных филаментов клеток кожи
- Экспрессия генов кератина при дифференцировке эпидермиса
- Функции кератина в эпидермисе и коже в целом
- Патология кератина как причина кожных болезней
- Роговой конверт эпидермиса - состав, функции
- Защитная и барьерная функция кожи как органа
- Строение и защитные функции рогового слоя эпидермиса кожи
- Липиды рогового слоя эпидермиса кожи и его барьерная функция