
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Экспрессия генов кератина при дифференцировке эпидермиса
Кожа служит ярким примером тесного взаимодействия между регулированием генов кератина и дифференцировкой эпидермиса. Более половины всех известных генов кератина экспрессированы исключительно в кожной ткани зрелых млекопитающих. Сложная структура эпителия взрослой кожи является результатом пространственно-временного регулирования экспрессии генов кератина и ряда других генов, связанных с дифференцировкой эпителия.
В клинической практике типирование кератина часто применяется для диагностики вида рака, определения стадии его дифференцировки (что позволяет прогнозировать дальнейшее развитие болезни), а также для идентификации происхождения клеток, образующих метастатические очаги. Данный подход применим не только для онкологических заболеваний (см. ниже).
В «тонком» межфолликулярном эпидермисе (например, в эпидермисе туловища) митотически активные клетки базального слоя выполняют функцию прогениторных клеток и постоянно экспрессируют основную пару кератинов К5 и К14 и в меньшей степени — К15. Начальному этапу дифференцировки соответствует появление пары К1/К10 вследствие мощной транскрипционной индукции, осуществляющейся за счет генов К5/К14, экспрессия которых понижается. Таким образом, кератины К1/К10 без труда обнаруживаются в самом нижнем супрабазальном слое эпидермиса.
Появление К1 и К10 взаимосвязано с резким изменением в порядке организации промежуточных филаментов кератина, которые в этот момент предстают в виде пучков. Еще один ген кератина типа II — К2—экспрессируется на более поздней стадии дифференцировки (например, в гранулярном слое).
Эпидермис ладоней и подошв приспособлен для того, чтобы выдерживать значительную механическую нагрузку, и здесь его толщина существенно больше, нежели на других участках кожи. Подобная функциональная особенность эпидермиса сказывается как на его структуре — он состоит из чередующихся первичных и вторичных гребешков, — так и на экспрессии кератинов в его клетках.
В толстых, испытывающих нагрузку первичных гребешках, основной дифференцировочный кератин К9 (тип I) способствует формированию более эластичного цитоскелета. В более тонких вторичных гребешках экспрессированы преимущественно постмитотические кератиноциты Кба (тип II) и К16 и К17 (тип I). По всей вероятности, кератины К16 и К17, сопоставимые по своим свойствам с К1, К9 и К10, обеспечивают здесь большую пластичность, создавая гибкие «шарнирные» регионы между более жесткими, богатыми кератинами К1/К9 первичными гребешками.
Описанная модель еще нуждается в экспериментальном подтверждении, однако косвенным ее доказательством служит увеличение экспрессии К6а, K6b, К16 и К17 в кератиноцитах, рекрутируемых с краев раны для участия в восстановлении поврежденного эпидермального барьера.
Эпидермальные заболевания часто сопровождаются нарушениями нормальной дифференцировки и, что неудивительно, практически всегда сопряжены с изменением в экспрессии генов кератина. Например, К6а, К6b и/или К17, в норме предназначенные для восстановления раневых поверхностей в эпидермисе туловища, эктопически задействованы в генезе псориаза и сходных гиперпролиферативных заболеваний, немеланоцитарного рака кожи, вирусных инфекций и других состояний, сопровождающихся воспалительным процессом. Сходное «замещение» К1 и К10 кератинов на К6, К16 и К17 возникает, если нормальные кератиноциты человека помещаются в культуру.
На ранних этапах злокачественного процесса при плоскоклеточном раке кожи отмечается прогрессивная редукция экспрессии К6/К16 при сохранности некоторой степени экспрессии К1/К10, что отражает достаточную степень дифференцировки, тогда как в дальнейшем экспрессия К1/К10 полностью прекращается при сохранности синтеза простых эпителиальных кератинов К8/К18, что указывает на низкий уровень дифференцировки и более агрессивное течение процесса. Эти вариации экспрессии генов кератина, вероятно, в значительной степени влияют на биологические свойства кератиноцитов.
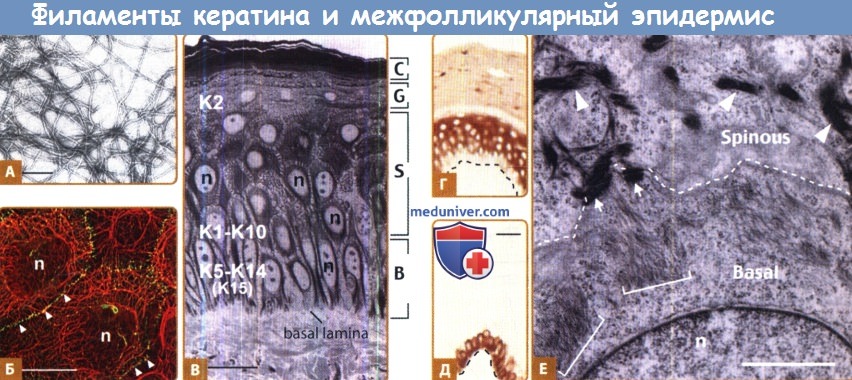
А. Филаменты, восстановленные in vitro из очищенных кератинов человека К5 и KI4. Визуализация методами негативного окрашивания и электронной микроскопии. (Бар = 150 нм.)
Б. Двойноемечение кератина (красныйхромофор) и десмоплакина, являющегося десмосомным компонентом (зеленый хромофор), методом непрямой иммунофлуоресценции культуры клеток эпидермиса человека.
Промежуточные филаменты кератина организованы в сеть, охватывающую всю цитоплазму, и прикреплены к десмосомальным участкам межклеточных контактов (обозначены стрелками) (Бар = 50μм.)
В. Залитый в смолу гистологический срез эпидермиса, взятого с туловища человека.
Видны базальный (В - basal), шиповатый (S - spinous), зернистый (G - granular) и роговой (С - cornified) клеточные слои (Бар = 50μм.)
Г и Д. Характерное распределение эпитопов кератина, наблюдаемое на гистологических срезах кожи человека.
Визуализация методом обнаружения антител. Г. К10 в основном сконцентрирован в дифференцирующихся супрабазальных слоях эпидермиса. Пунктирной линией обозначена базальная пластинка.
Д. К14 находится в базальном слое, где расположены эпидермальные прогениторные клетки (Бар = 50μм.)
Е. Стандартная трансмиссионная электронная микроскопия пограничной зоны между базальными и супрабазальными клетками в эпидермисе туловища мыши.
Образец материала, на микрофотографии, ориентирован так же, как и на рис. В.
Организация кератиновых филаментов в форме рыхлых пучков (обозначены квадратными скобками) коррелирует с экспрессией К5-К14 в базальных клетках,
в то время как образование более густых электронно-плотных пучков филаментов (обозначены стрелками) соответствует начальной фазе экспрессии К1-К10 в рано дифференцирующихся клетках.
Стрелки указывают на десмосомы, соединяющие две клетки (Бар = 2μм.) n = ядро.
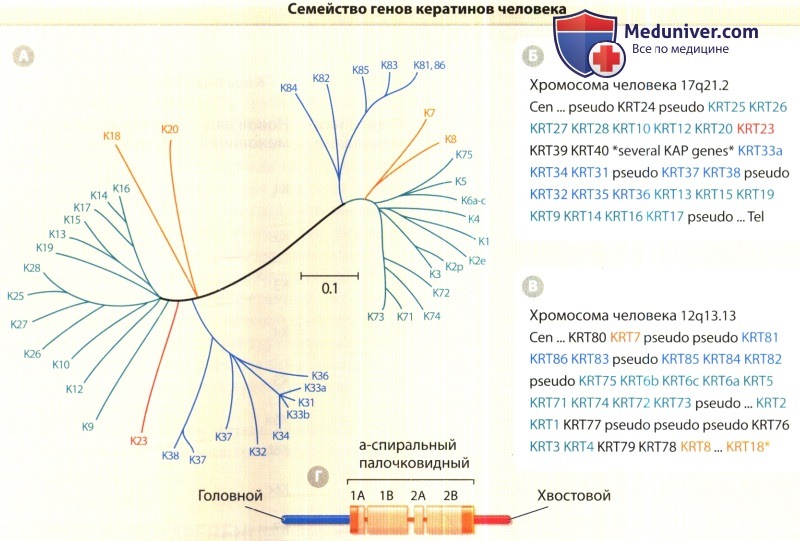
А. Сравнительная схема базовой структуры кератинов человека, построенная с применением компьютерных программ ClustaW и TreeView.
Связанность последовательностей обратно коррелирует с длиной линий, соединяющих различные последовательности, а также с количеством и месторасположением точек ответвления.
Для сравнения взяты последовательности головного и центрального палочковидных доменов каждого кератина.
Для большей ясности некоторые кератины в данной схеме не указываются. «Дерево» имеет два главных ответвления, соответствующих последовательностям кератинов типов I и II.
Каждая из частей этой ведущей дихотомии, в свою очередь, подразделяется на основные подгруппы (они обозначены разными цветами).
Б. Организация генов кератинов функционального типа I, сгруппированных у человека на хромосоме 17.
Исключением является К18 (помечен звездочкой в пункте В), находящийся на участке, граничащем с теломерой (Tel) группы генов типа II.
Большое число генов, кодирующих ассоциированные с кератинами протеины (КАР), прерывает группу генов типа I между KRT40 и KRT33A.
В. Организация функциональных генов кератинов типа II, сгруппированных у человека на хромосоме 12. Гены К8 и К18 разделяются 450000 пар оснований.
Г. Схематическое изображение трехчастной структуры домена, общей для всех кератинов и других белков промежуточных филаментов.
Центральный а-спиральный палочковидный домен выступает в качестве основного детерминанта самосборки.
Он разделен на субдомены 1 А, 1 В, 2А и 2В и фланкирован неспиральными «головным» и «хвостовым» доменами — N-концевым и С-концевым соответственно.
Оба конца палочковидного домена содержат от 15 до 20 участков аминокислот (обозначены красным цветом), в высокой степени консервативных у всех белков промежуточных филаментов Cen = центромера;
several КАР genes = несколько генов, кодирующих ассоциированные с кератинами протеины; pseudo = псевдоген (нефункциональный); tel = теломера.
- Рекомендуем далее ознакомиться со статьей "Функции кератина в эпидермисе и коже в целом"
Оглавление темы "Гистология, физиология кожи.":- Клеточная и генная терапия стволовыми клетками эпидермиса
- Классификация кератинов человека
- Кератиновые белки промежуточных филаментов клеток кожи
- Экспрессия генов кератина при дифференцировке эпидермиса
- Функции кератина в эпидермисе и коже в целом
- Патология кератина как причина кожных болезней
- Роговой конверт эпидермиса - состав, функции
- Защитная и барьерная функция кожи как органа
- Строение и защитные функции рогового слоя эпидермиса кожи
- Липиды рогового слоя эпидермиса кожи и его барьерная функция