
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Липиды рогового слоя эпидермиса кожи и его барьерная функция
Сканирование с помощью конфокального лазерного микроскопа и рентгеновский микроанализ показали, что основной путь проникновения веществ сквозь кожу представляет собой извилистую траекторию между корнеоцитами, что подтверждает незаменимую роль межклеточных липидов в регулировке барьерной функции. К основным классам липидов рогового слоя относятся холестерин, свободны жирные кислоты и церамиды.
а) Холестерин. Холестерин — вероятно, наиболее распространенный липид в организме в целом — является составной частью плазмалеммы и межклеточных липидных пластинок рогового слоя. Хотя базальные клетки способны поглощать холестерин из циркулирующей крови, в основном холестерин синтезируется из ацетата in situ.
Эпидермальные кератиноциты — основной тип клеток эпидермиса — активно синтезируют несколько липидов, в том числе холестерин и свободные жирные кислоты. Катализатором стадии, ограничивающей скорость биосинтеза холестерина, является гидроксиметилглютарил-коэнзим А-редуктаза. Синтез эпидермального холестерина регулируется данными ферментами, и его скорость повышается во время восстановления барьера проницаемости.
б) Свободные жирные кислоты. Кожа содержит свободные жирные кислоты, а также жирные кислоты, связанные в триглицериды, фосфолипиды, гликозилцерамиды и церамиды. Длина цепочки свободных жирных кислот в эпидермисе варьирует от 12 до 24 атомов углерода. Ограничивающие скорость синтеза ферменты — ацетил-СоА карбоксилаза и синтаза жирной кислоты — присутствуют в эпидермисе в значительной мере автономно.
Насыщенные и мононенасыщенные жирные кислоты, в отличие от диненасыщенных и полиненасыщенных кислот, синтезируются в эпидермисе. Номенклатура жирных кислот определяется положением первой (отсчитывая от терминальной метиловой группы) двойной связи в молекуле. Эссенциальные ω-6-ненасыщенные кислоты должны поступать в организм с пищей и через кровообращение, они могут быть также введены при местной терапии. Неэссенциальная мононенасыщенная жирная кислота—олеиновая кислота—представляет собой ω-9 жирную кислоту.
Линолевая кислота — наиболее важная двойная ненасыщенная жирная кислота — является ω-6 кислотой. Большое значение имеет также и а-линолевая кислота (ω-3). Изменения кожи, обусловленные дефицитом ω-3 жирных кислот, на данный момент не известны; однако предполагается, что со-3 жирные кислоты необходимы для разрешения воспалительного процесса, ω-3 жирные кислоты содержатся в рыбе, источником ω-6 жирных кислот являются растительные масла. Дефицит эссенциальных жирных кислот, возникающий вследствие изменений в рационе питания или в результате мальабсорбции, приводит к синдрому дефицита эссенциальных жирных кислот, для которого характерны значительные изменения в эпителии, включая эпидермис, что было подтверждено в экспериментах на крысахи мышах.
В подобном состоянии эпидермис становится шершавым, шелушащимся и гиперемированным, а проницаемость серьезно нарушается. Кроме того, при дефиците эссенциальных жирных кислот могут наблюдаться тяжелая бактериальная инфекция, замедленное заживление ран и алопеция. Линолевая кислота входит в состав фосфолипидов, глюкозилцерамидов, церамида 1, церамида 4 и церамида 9. Предполагается, что метаболит линолевой кислоты — γ-линолевая кислота — играет важную роль при атопической экземе.
в) Церамиды. Церамид представляет собой соединенную амидной связью жирную кислоту, содержащую длинноцепочечный аминоспирт, который называется сфингоидной основой. Длины карбоновых цепей, соединенных амидной связью жирных кислот и сфингоидных основ, в тканях большинства млекопитающих составляют от 16 до 26 и от 18 до 20 атомов углерода соответственно. Хотя сфинголипиды, включая гликосфинголипиды и фосфосфинголипиды, представлены во всех тканях млекопитающих, также описано и специфическое распределение этих молекул в различных тканях.
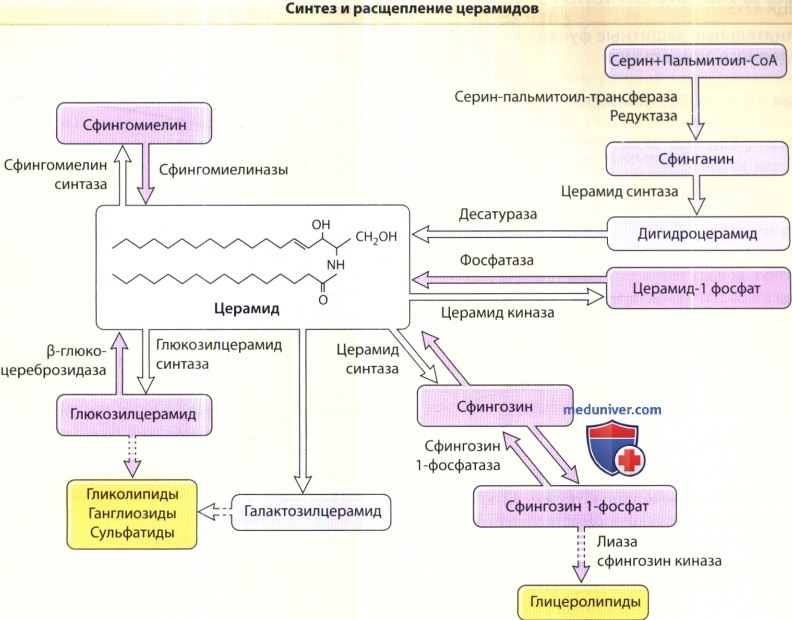
Эпидермис и селезенка богаты глюкозилцерамидом. Галактозилцерамид, отсутствующий в кератиноцитах, присутствует в головном мозге. В роговом слое церамид является основным липидным компонентом, и его вес здесь может достигать 30-40% от веса всех липидов, в то время как в других тканях млекопитающих он составляет менее 10% от веса холестерина или фосфолипидов. Столь высокое содержание церамидов не наблюдается ни в зернистом, ни в шиповатом, ни в базальном слоях эпидермиса. Данный факт свидетельствует о том, что основным условием накопления церамидов является терминальная дифференцировка.
Роговой слой содержит как минимум девять различных церамидов. Кроме того, существуют два белково-связанных церамида — церамид А и церамид В. Данные церамиды ковалентно связаны с белками рогового конверта, главным образом — с инволюкрином.
Церамиды синтезируются серин-пальмитоил-трансферазой — ферментом, лимитирующим скорость,— а также гидролизом глюкозилцерамида (посредством β-глюкоцереброзидазы) и сфингомиелина (посредством кислотной сфингомиелиназы). Если серин-палмитоил-трансферазы и β-глюкоцереброзидаза осуществляют синтез всех видов церамидов, то из сфингомиелиназы синтезируются только церамид 2 и церамид 5, так как сфингомиелин содержит негидроксильные кислоты.
Транспорт липидов. Для поддержания проницаемости кожного барьера кератиноцитам необходимо большое количество холестерина. АВСА1 —мембранный транспортный белок, ответственный как за приток холестерина, так и за регулирование уровня холестерина в клетках.
Исследования показали, что АВСА1 экспрессирован в культуре кератиноцитов человека и в эпидермисе мыши. Экспрессия АВСА1 в культуре кератиноцитов усиливается за счет активации нескольких рецепторов: Х-рецептора печени, рецепторов, активирующих пролиферацию пероксисом,— (PPAR)-a, PPAR-ss/6 и ретиноидного Х-рецептора. Таким образом, в регулировании уровня холестерина, необходимого для осуществления функции проницаемости кожного барьера, участвуют АВСА1, Х-рецептор печени и рецепторы PPAR. Транспорт и метаболизм жирных кислот в клетке регулируются белками, связанными с жирными кислотами.
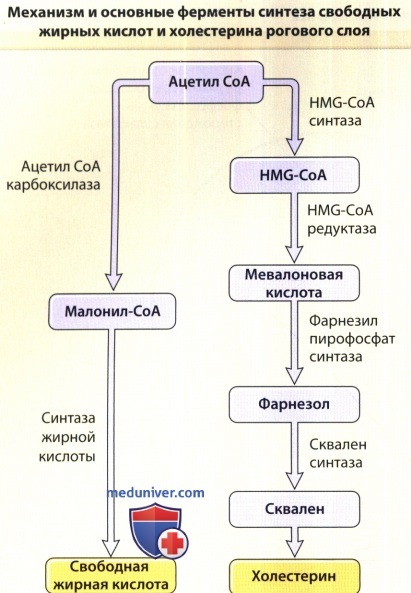
СоА = Коэнзим A; HMG-CoA = Гидроксиметилглютарил СоА.
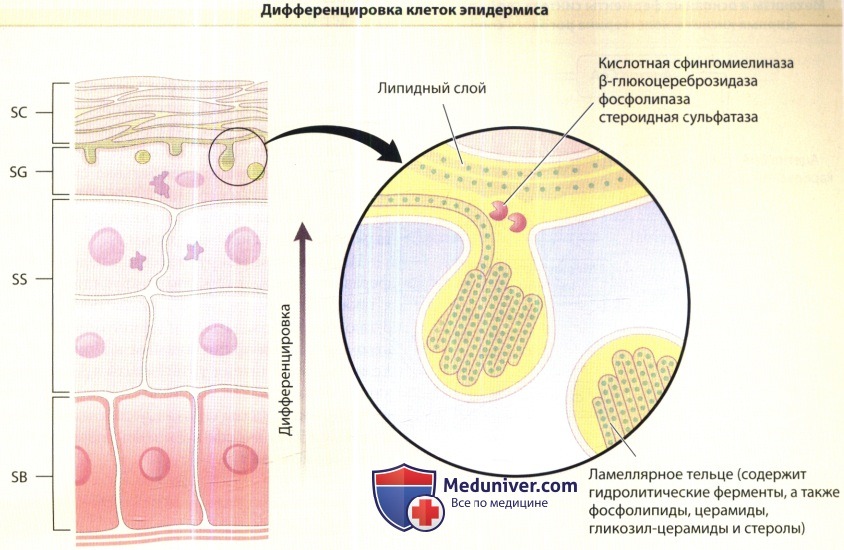
которые содержат ранее сформировавшиеся липидные структуры и гидролитические ферменты.
Их содержимое вытесняется в пограничную зону между зернистым (SG) и роговым (SC) слоями и подвергается значительной трансформации.
SB = базальный слой.
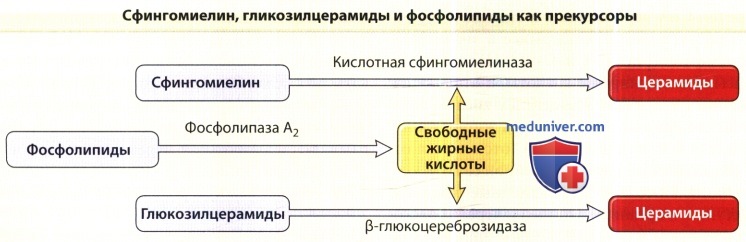
фосфолипиды прекурсорами синтеза жирных кислот.
- Вернуться в оглавление раздела "дерматология."
Оглавление темы "Гистология, физиология кожи.":- Клеточная и генная терапия стволовыми клетками эпидермиса
- Классификация кератинов человека
- Кератиновые белки промежуточных филаментов клеток кожи
- Экспрессия генов кератина при дифференцировке эпидермиса
- Функции кератина в эпидермисе и коже в целом
- Патология кератина как причина кожных болезней
- Роговой конверт эпидермиса - состав, функции
- Защитная и барьерная функция кожи как органа
- Строение и защитные функции рогового слоя эпидермиса кожи
- Липиды рогового слоя эпидермиса кожи и его барьерная функция