
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Регуляция клеточных реакций на повреждение ДНК
Сохранению целостности генома способствуют несколько клеточных реакций в ответ на повреждение ДНК, в том числе арест клеточного цикла, апоптоз (запрограммированная гибель клетки) и репарация ДНК. Эти реакции требуют точной регуляции, в передаче сигнала о повреждении ДНК и регуляции реакций на повреждение ДНК участвует множество белков.
Для восстановления разных типов повреждения ДНК, вызванных различными поражающими факторами, требуются различные ответные реакции. Упрощенная версия этого сложного механизма представлена на рисунке ниже.
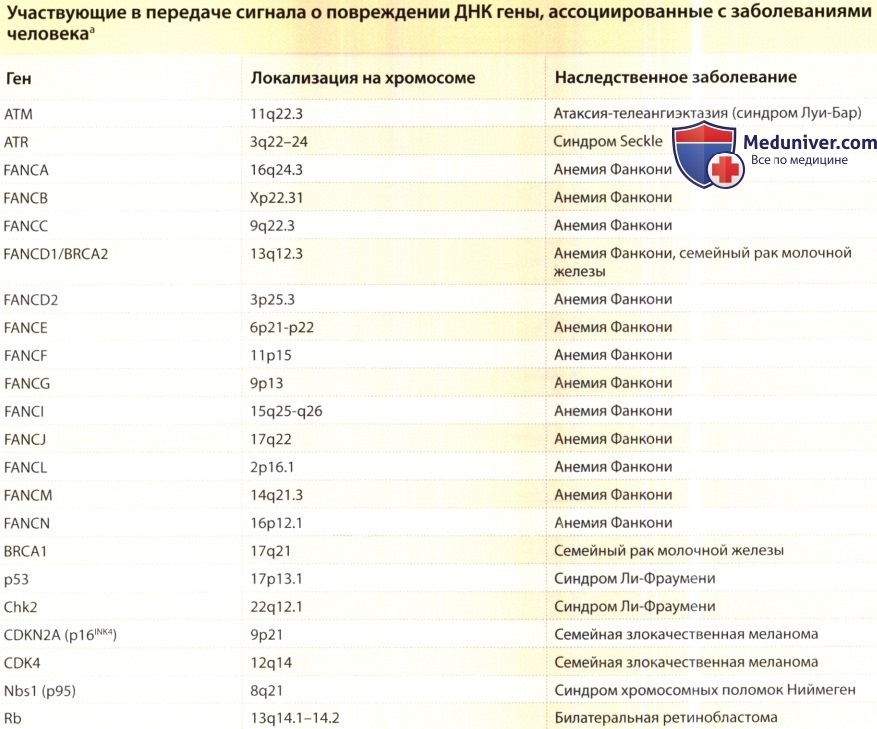
Как и при дефектах генов, кодирующих репарацию ДНК, дефекты многих из этих генов, кодирующих передачу сигнала о повреждении ДНК, также участвуют в развитии наследуемой нестабильности генома.
Для таких заболеваний характерен повышенный риск рака вследствие нестабильности генома в результате дефектной сигнализации о повреждении ДНК.
Ген супрессора образования злокачественных опухолей р53, названный «стражем генома», играет ключевую роль в регуляции и контроле этих реакций, и его мутация обнаруживается при ряде злокачественных опухолей, включая плоскоклеточный рак кожи.
Основными регуляторами р53 при развитии клеточных реакций на повреждение ДНК являются гены АТМ (атаксии телеангиоэктазии мутировавший) и ATR (связанный с атаксией телеангиоэктазией и Rad3). Одна из множества функций р53 — регуляция клеточного цикла в ответ на повреждение ДНК.
После деления (митоза) клетки содержат 23 пары хромосом и вступают в G1 фазу клеточного цикла.
Затем хромосомы реплицируются в процессе этапа синтеза ДНК, или S-фазы, в результате чего количество хромосом удваивается (фаза G2), после чего клетка вступает в фазу митоза (фаза М). В ответ на повреждение клетка может остановить («арестовать») свой цикл на его специфических этапах, которые называются контрольными точками цикла.
Важным нисходящим эффектором при предотвращении вступления клетки в S-фазу клеточного цикла (контрольная точка G1/S) является р21. Кроме того, р53 индуцирует репарацию с удалением нуклеотида путем транскрипционной индукции ХРС,ХРЕ/р48 и GADD45. Это указывает на то, что в ответ на повреждение ДНК способность клетки к репарации поврежденной ДНК может усиливаться.
Если клетки вступают в S-фазу с невосстановленым повреждением ДНК, или если клетки подверглись действию УФ во время S-фазы, обычная ДНК-полимераза останавливается на фотопродуктах ДНК и открепляется от цепочки ДНК.
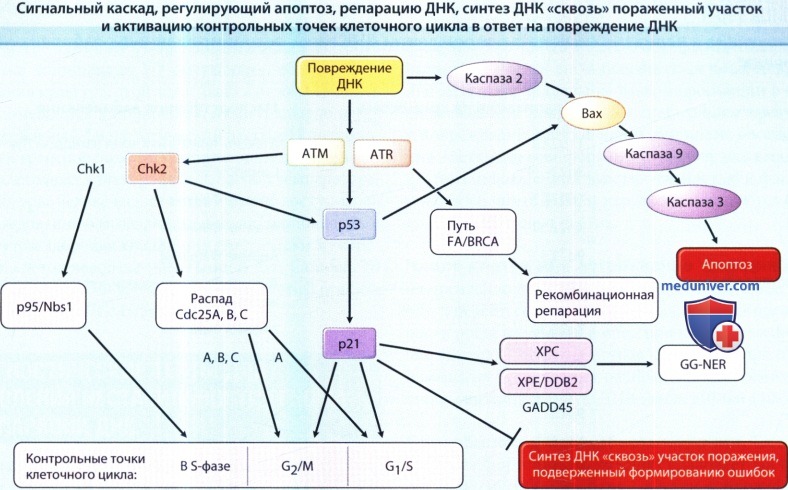
Это сильно упрощенная диаграмма, на которой изображены только самые важные участники сложных и переплетенных сетей передачи сигнала о повреждении ДНК.
Традиционно считается, что ATM (ген атаксии телеангиоэктазии мутировавший) активируется (фосфорилируется) в ответ на ионизирующее излучение (ИИ),
a ATR (ген атаксии телеангиоэктазии и Rad3) активируется ультрафиолетовым (УФ) излучением, однако последние данные указывают на то, что они оба активируются как ИИ, так и УФ.
ATM/ATR могут активировать (т.е. фосфорилировать) р53 как прямо, так и косвенно посредством активации (фосфорилирования) Chk2 (мишень ATM) или Chk1 (мишень ATR).
Активированный р53 активирует контрольную точку G1/S (т.е. «арестовывает» клетки в фазе G1) путем транскрипционной активации р21 и последующего ингибирования циклин-зависимых киназ,
которые обычно переводят клетки из фазы G1 в фазу S клеточного цикла.
Контрольная точка G1/S также активируется индуцированным Chk1/Chk2 фосфорилированием с распадом Cdc25A, что в результате приводит к невозможности активации циклин-зависимых киназ.
Фосфорилирование с последующим распадом Cdc25A, Cdc25B и Cdc25C также опосредует «арест» в G/М, как и р21. «Арест» клетки в S-фазе опосредован активацией (фосфорилированием) p95/Nbs1.
Р53 также индуцирует общую геномную репарацию с удалением нуклеотида (GG-NER) посредством транскрипционной активации ХРЕ/р48 и GADD45.
Было показано, что синтез ДНК «сквозь» пораженные участки тормозится р53 при участии р21.
Рекомбинационная репарация опосредуется метаболическим путем, участвующим в развитии анемии Фанкони (FAJ/BRCA, который, в свою очередь, зависит от активации ATR.
Митохондриальный путь развития апоптоза инициируется путем активации Вах каспазой 2 и р53, инициаторной каспазой 9 и эффекторными каспазами 3, 6 и 7.
Продукты генов, имеющих дефект при наследственных заболеваниях человека, обведены рамкой.
DDB2—протеин 2, связывающий повреждение ДНК; ХРС, ХРЕ—комплементационные группы С и Е при пигментной ксеродерме.
Для таких ситуаций клетка оснащена различными специальными ДНК-полимеразами для синтеза ДНК в том числе в поврежденном участке.
Одной из них является ДНК-полимераза; она способна обходить фотопродукты ДНК, но в ходе такой транскрипции может приводить к формированию мутаций.
Она мутирует при одном из вариантов пигментной ксеродермы, который клинически от пигментной ксеродермы неотличим. Это указывает на большое значение этой защиты второй линии от мутагенных и канцерогенных последствий формирования фотопродуктов ДНК.
Сообщалось, что р53 и р21 также снижают активность такого синтеза ДНК «сквозь» поврежденные участки с целью снижения мутагенной активности за счет уменьшения обходов поврежденного участка при транскрипции.
Если не удается провести синтез ДНК «сквозь» участок поражения, клетка может использовать рекомбинационную репарацию для устранения остановки репликации второй нити.
При вовлечении в ответную реакцию на повреждение УФ-лучами эта третья линия защиты опосредуется активацией метаболического пути, участвующего в развитии реакции на повреждение ДНК при анемии Фалькони/BRCA. Точный механизм, инициирующий эти сигнальные каскады реакции на повреждение ДНК, изучается.
Считается, что теломеры, которые являются повторами TTAGGG и завершают концы хромосом, формируют структуру петли, играя важную роль в детекции повреждения ДНК, например, путем открытия петли теломера.
В качестве других сенсоров могут выступать остановившиеся ДНК- или РНК-полимеразы или белки, выявляющие изгибание спирали ДНК в месте повреждения ДНК.
Апоптоз представляет собой регулируемый физиологический процесс, приводящий к гибели клеток, характеризующийся уменьшением клетки, везикуляцией мембраны и фрагментацией ДНК. Елавными регуляторами апоптоза являются группа цистеиновых протеаз, названных «каспазами».
В качестве триггеров могут выступать как внешние, так и внутренние по отношению к клетке факторы (например, повреждение ДНК), которые вовлекают только различные инициаторные каспазы (например, в ответ на повреждение ДНК каспазу-2), но и общие эффекторные каспазы.
- Рекомендуем далее ознакомиться со статьей "Симптомы нестабильности генома и диагностика нарушения репарации ДНК"
Оглавление темы "Канцерогенез рака кожи.":- Канцерогенез рака кожи. Факторы риска рака кожи
- Повреждение ДНК кожи и ее репарация. Гены репарации ДНК
- Регуляция клеточных реакций на повреждение ДНК
- Симптомы нестабильности генома и диагностика нарушения репарации ДНК
- Химический канцерогенез рака кожи. Окружающая среда как причина рака
- Онкогены, гены-супрессоры роста опухоли и наследственные причины рака
- Модели рака кожи на животных для изучения онкогенеза