
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Рецепторы комплемента и их лиганды
• Ряд важных функций комплемента опосредуются связыванием комплементарных белков с рецепторами. Рецептор CR1 задействован в фагоцитозе, удалении иммунных комплексов и в механизме ослабления или подавления ответа на стимул. CR2 является важным стимулятором гуморального иммунитета. С3 и С4 способствуют фагоцитозу.
• Регуляция активации комплемента осуществляется определенными белками сыворотки и клеточной поверхности. Многие из регуляторных белков клеточной поверхности, экспрессированные на клетках человека, не экспрессированы на микробах, что защищает клетки человека от повреждения комплементом.
• Некоторые из регуляторных белков понижают активность комплемента, вытесняя компоненты начальных ступеней каскада.
К ним относятся ингибитор С1, фактор Н, ингибитор связывания С4, фактор, ускоряющий диссоциацию, мембранный кофакторный белок (МСР), а также рецептор CR1.
Существует ряд белков, взаимодействующих с комплементом и служащих для опосредования или регулировки его функций. С5а—рецептор, который является членом семейства семи-а-спиральных трансмембранных рецепторов, сопряженных с G-белком—был отмечен в предыдущем разделе. К числу наиболее подробно описанных рецепторов комплемента относятся CR1-CR4
Рецептор комплемента типа 1 — CR1 (CD35)—-принадлежит к семейству белков, называемых регуляторами активации комплемента. Они имеют общую структуру, состоящую из множества коротких консенсусных повторов, известных также как белковые повторы контроля комплемента. CR1 связывает С3b с С4b и экспрессирован на клетках периферической крови, включая моноциты, В- и Т-лимфоциты, нейтрофилы, эозинофилы и эритроциты, фолликулярные дендритные клетки и кератиноциты.
На фагоцитарных клетках связывание CR1 с С3b и С4b приводит к фагоцитозу частиц, опсонизированных фрагментами комплемента, а также к активации бактерицидного механизма в фагоцитарных клетках. На эритроцитах связывание CR1 иммунными комплексами, покрытыми С3b или С4b, вызывает перемещение комплексов в селезенку и печень, где они удаляются из кровообращения фагоцитами. Таким образом, CRI служит важным медиатором функции комплемента.
CR1 задействован в диссоциации комплексов С3-конвертазы, в связи с чем он также может использоваться для подавления активации комплемента.
Рецептор комплемента типа 2 — CR2 (CD21) — также относится к белковому семейству регуляторов активации комплемента. CR2 связывает фрагменты комплемента С3 — iC3b, C3dg и C3d, и, кроме того, вирус Эпштейна-Барр, интерферон-а и иммунорегуляторный белок CD23. CR2 экспрессирован на субпопуляциях В- и Т-лимфоцитов, базофилах, тучных клетках, фолликулярных дендритных клетках и некоторых эпителиальных клетках. На В-клетках CR2 служит ко-рецептором их активации. Если CR2 связывается посредством C3d, уровень активации В-клеток возрастает на несколько порядков.
На дендритных клетках взаимодействие с CR2 приводит к захвату иммунных комплексов в зародышевых центрах. По всей видимости, CR2 также играет определенную роль в презентации антигенов Т-клеткам.
Рецептор комплемента типа 3 — CR3 (CD11b/CD18, Mac-1) — является молекулой клеточной поверхности интегрина. Данная молекула экспрессируется на моноцитах, нейтрофилах, ЕК-клетках и тучных клетках. Функция этого рецептора состоит в содействии фагоцитозу микробов посредством связывания с iC3d и прямого связывания с микробами. Рецептор типа 3 взаимодействует с молекулой межклеточной адгезии 1, которая эндогенно эксрессирована на эндотелиальных клетках для стабилизации адгезии лейкоцитов к эндотелию, способствуя привлечению лейкоцитов из кровообращения в ткань.
Рецептор комплемента типа 4 — CR4 (CD11с/CD18) — также представляет собой молекулу клеточной поверхности интегрина. Он экспрессирован на моноцитах, нейтрофилах, ЕК-клетках и дендритных клетках, и его функция, вероятно, сходна с функцией CR3.
Среди недавно описанных рецепторов комплемента отметим SIGN-R1, который связывает C1q и вырабатывается макрофагами краевой зоны селезенки, а также CRIg (рецептор комплемента из семейства иммуноглобулинов), который связывает С3b и iC3b и экспрессируется на субпопуляции тканевых макрофагов.
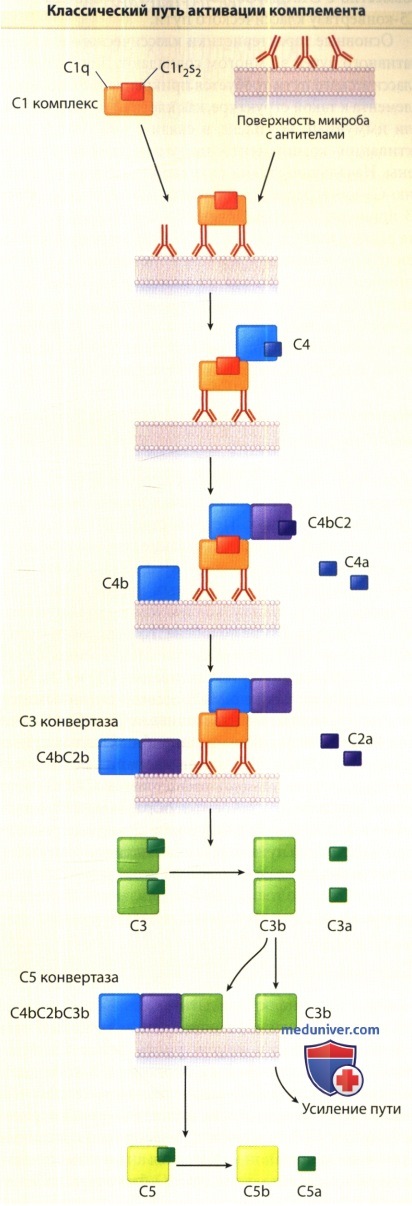
Показаны стадии активации от начального прикрепления C1q к комплексу антиген-антитело посредством расщепления С5 на С5а и С5b.
Завершающие стадии активации комплемента представлены на рисунке ниже.
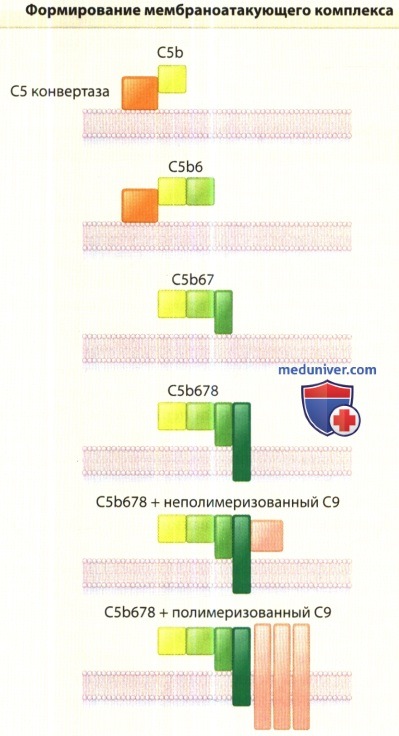
Альтернативный, классический и лектиновый пути сходятся на этапе образования С5b и его последующего соединения с компонентами С6, С7, С8 и С9.
Полимеризация С9 приводит к возникновению трубчатых структур на клеточной мембране.
- Рекомендуем далее ознакомиться со статьей "Регуляция активации комплемента"
Оглавление темы "Гуморальный иммунитет при болезнях кожи.":- Активация и связывание антигена В-лимфоцитами
- Роль В-лимфоцитов в развитии кожных болезней
- Этапы альтернативного пути активации комплемента
- Этапы классического пути активации комплемента
- Этапы лектинового пути активации комплемента
- Общие завершающие этапы активации комплемента
- Функции белков комплемента
- Рецепторы комплемента и их лиганды
- Регуляция активации комплемента
- Наследственные нарушения в системе комплемента
- Участие комплемента в развитии болезней кожи