
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
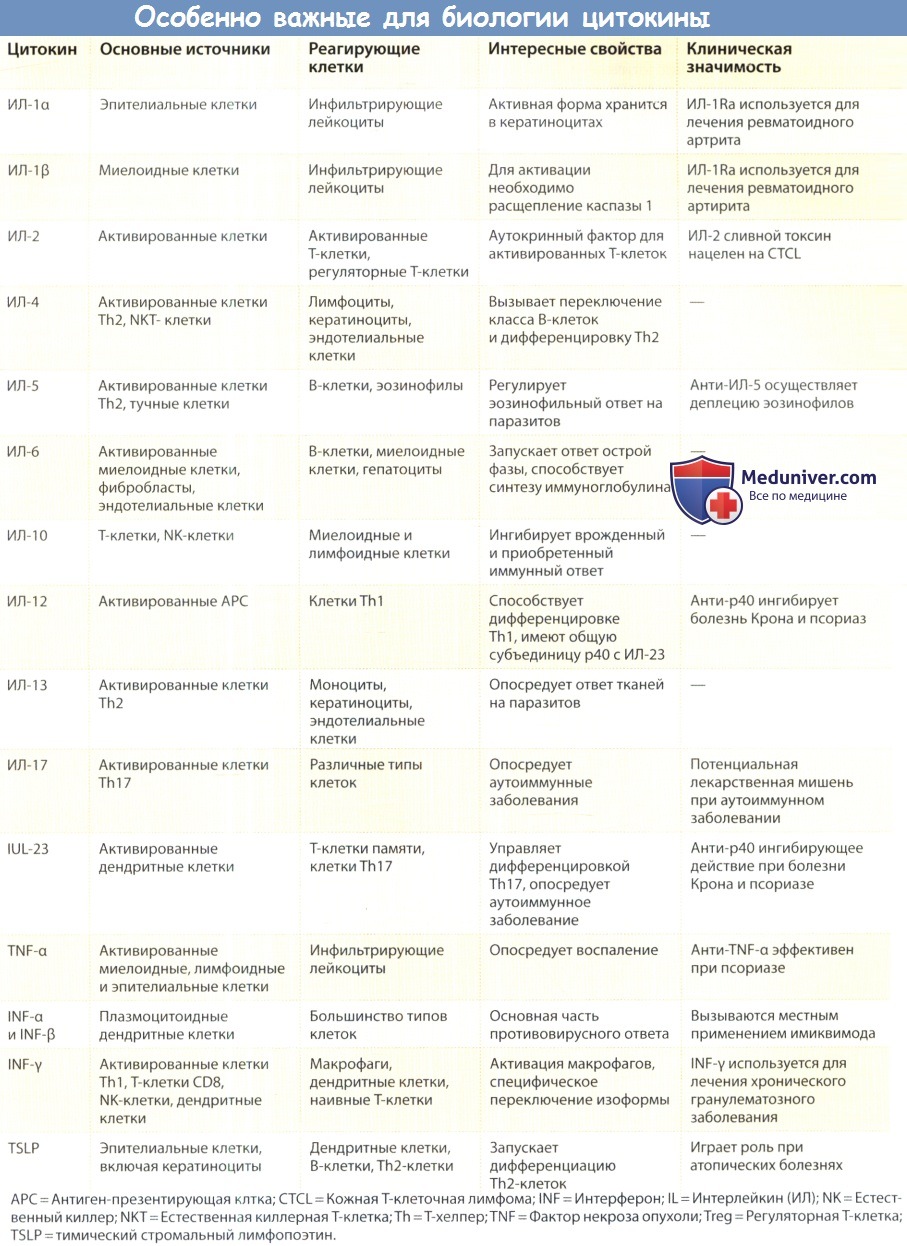
Цитокины интерлейкин-4 (ИЛ-4) и интерлейкин-13 (ИЛ-13)
ИЛ-4 и ИЛ-13 являются продуктами активированных клеток Th2 с ограниченной структурной гомологией (примерно 30%) и перекрестными, но различными биологическими действиями. Специфический рецептор для ИЛ-4, который не связывает ИЛ-13, обнаружен на Т-клетках и ЕК-клетках.
Он состоит из ИЛ-4Rа (CD124) и ус и передает сигналы через Jak1 и Jak3. Комплекс рецепторов, который может связывать либо ИЛ-4, либо ИЛ-13, обнаружен на кератиноцитах, эндотелиальных клетках и других негематопоэтических клетках. Он состоит из ИЛ-13Rа1 (CD213al) и ИЛ-4Rа и передает сигналы через Jak1 и Jak2. Эти рецепторы экспрессированы на низких уровнях в покоящихся клетках, и их экспрессию усиливают различные активирующие сигналы.
Интересно, что воздействие моноцитов на ИЛ-4 или ИЛ-13 подавляет экспрессию ИЛ-4Rа и ИЛ-13Rа1, в то время как в кератиноцитах наблюдается противоположный эффект. Оба пути передачи сигнала при активации STAT6 сливаются, что необходимо и достаточно для запуска дифференцировки Th2. ИЛ-13Rа2 является рецептором, расположенным на поверхности клетки. Он гомологичен ИЛ-13Rа1 рецептору, который специфически связывает ИЛ-13. Относительно последнего точно неизвестно, передает ли он какие-либо сигналы.
Биологические эффекты рецептора ИЛ-4 различаются в зависимости от специализации клеток, но чаще всего сводятся к его принципиальной роли как фактора роста и дифференцировки клеток Th2. Воздействие ИЛ-4 на наивные Т-клетки стимулирует их к пролиферации и дифференцировке в клетки Th2, которые синтезируют больше ИЛ-4, что, в свою очередь, приводит к аутокринной стимуляции, которая удлиняет реакции по типу Th2.
Так, экспрессия ИЛ-4 на раннем этапе иммунного ответа может инициировать каскад развития клеток Th2, что приводит к превалированию ответа по типу Th2. Гены, кодирующие ИЛ-4 и ИЛ-13, расположены в кластере с ИЛ-5, который во время дифференцировки Th2 подвергается структурным изменениям, которые ассоциируются с усилением экспрессии. Хотя при активации синтез ИЛ-4 наивными Т-клетками может снижаться, ИЛ-4 также вырабатывается активированными естественными киллерными Т-клетками.
Тучные клетки и базофилы также выделяют предварительно синтезированный ИЛ-4 из секреторных гранул в ответ на опосредуемые FcεRl сигналы. Важная активность ИЛ-4 — это стимуляция переключения класса генов иммуноглобулина в В-клетках. Нуоциты и естественные хелперы представляют собой недавно открытые популяции клеток, участвующие в формировании врожденного иммунитета. Они являются ранним источником ИЛ-13 при глистной инвазии.
Тh2, ИЛ-4 и ИЛ-13 являются медиаторами атопического иммунитета в качестве решающих факторов для дифференцировки и эффекторной функции. Дополнительно к контролю над поведением эффекторных клеток они также непосредственно воздействуют на резидентные клетки тканей, например при воспалительных реакциях в воздухоносных путях.
- Рекомендуем далее ознакомиться со статьей "Цитокины интерлейкин-9 (ИЛ-9) и интерлейкин-21 (ИЛ-21)"
Оглавление темы "Цитокины иммунологии кожи.":- Пути передачи сигнала цитокинами кожи
- Нуклеарный фактор кВ (NF-kB), ингибитор кВ и первичные цитокины кожи
- Участие белков JAT и STAT в пути сигнала
- Цитокины семейства интерлейкинов-1: ИЛ-1, ИЛ-18, ИЛ-33
- Цитокин фактор некроза опухоли (ФНО) и его рецепторы
- Цитокины семейства ИЛ-17
- Семейство рецепторов гемотопоэтина (семейство рецепторов хемокинов класса I)
- Цитокины интерлейкин-2 (ИЛ-2) и интерлейкин-15 (ИЛ-15)
- Цитокины интерлейкин-4 (ИЛ-4) и интерлейкин-13 (ИЛ-13)
- Цитокины интерлейкин-9 (ИЛ-9) и интерлейкин-21 (ИЛ-21)