
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Интерфероны как прототипы сигнализации через JAK/STAT
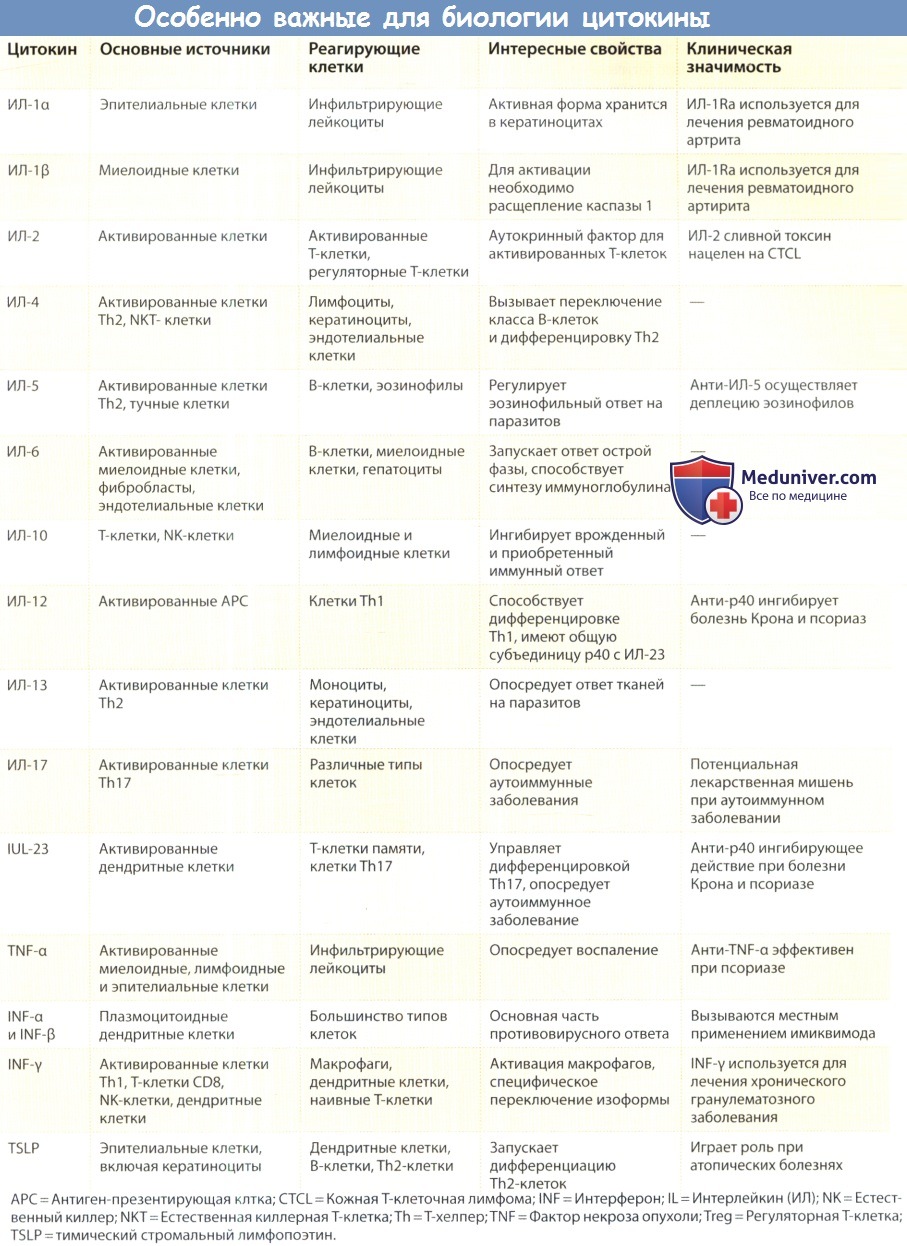
Интерфероны — одно из первых семейств цитокинов, которые были детально описаны. Вначале интерфероны подразделяли на три класса: (1) ИНФ-α (лейкоцитарные интерфероны), (2) ИНФ-β (фибробластные интерфероны) и (3) ИНФ-γ (иммунные интерфероны).
Интерфероны α и β называют общим термином «ИНФ типа 1», все эти молекулы сигнализируют через один и тот же двухцепочечный рецептор (рецептор ИНФ αβ). Второй ИНФ-рецептор представляет собой характерный двухцепочечный рецептор, специфичный для ИНФ-у. Оба эти ИНФ-рецептора присутствуют на многих типах клеток в коже, а также в других тканях.
Каждая из цепей, составляющих эти два ИНФ-рецептора, соединяется с одной из Jak-киназ (Тук2 и Jak1 для INFaβR и Jakl и Jak2 для 1NF у R). Эффективная передача сигнала после связывания ИНФ произойдет только в присутствии обеих цепей и двух функциональных Jak-киназ. Сейчас идентифицирован новый класс интерферонов, известный как ИНФ-А, или интерфероны типа III, которые имеют низкую степень гомологии с обоими интерферонами типа I и ИЛ-10.
К членам этого класса относятся ИЛ-28А, ИЛ-28В и ИЛ-29. Хотя эффекты этих цитокинов похожи на эффекты интерферонов типа I, они являются менее сильными. Такие интерфероны типа III используют общий рецептор, который состоит из β-цепи рецептора ИЛ-10 в связи с a-цепью рецептора ИЛ-28.
К стимулам, которые могут вызвать высвобождение из клеток интерферонов типа I, относятся вирусы, двухцепочечная ДНК и продукты бактерий. Плазмацитоидные дендритные клетки возникли как особенно мощный клеточный источник интерферонов типа I. Многие из эффектов ИНФ типа I прямо или косвенно повышают сопротивляемость организма распространению вирусной инфекции. Дополнительными эффектами, которые опосредуются ИНФ αβR, являются повышенная экспрессия молекул класса I главного комплекса гистосовместимости (МНС) и стимуляция активности ЕК-клеток. Это оказывает не только хорошо известный противовирусный эффект.
ИНФ-α может также модулировать Т-клеточные реакции, способствуя развитию Т-клеточного ответа по типу Th1. И, наконец, интерфероны типа I также ингибируют пролиферацию различных типов клеток, что служит обоснованием их применения в лечении различных злокачественных опухолей. Формы ИНФ-a находят широкое клиническое применение, в том числе при волосатоклеточном лейкозе, злокачественных поражениях кожи и инфицировании вирусом папилломы человека. Некоторые из заболеваний, которые реагируют на терапию ИНФ типа I, отвечают и на воздействие местных иммуномодулирующих агентов, таких как имиквимод.
Этот препарат синтетического имидазохинолина служит агонистом рецептора TLR7, естественным лигандом которого является одноцепочечная РНК. Стимуляция имиквимодом клеток, экспрессирующих TLR7, вызывает местное высвобождение больших количеств ИНФ типа I из плазмоцитоидных дендритных клеток, которые могут запустить антивирусный и противоопухолевый эффект при генитальных бородавках, поверхностной базальноклеточной карциноме и актинических кератозах. Резиквимод — родственный синтетический препарат, активирующий и TLR7, и TLR8, возбуждая несколько иной спектр цитокинов.
Выработка ИНФ-γ ограничена ЕК-клетками, Т-клет-ками CD8 и Т-клетками типа Тh1 CD4. Клетки Тh1 вырабатывают ИНФ-γ после активации Т-клеточного рецептора, а ИЛ-12 может обеспечить сильный костимулирующий сигнал для синтеза ИНФ-γ Т-клетками.
ЕК-клетки производят ИНФ-γ в ответ на цитокины, высвобождаемые макрофагами, в том числе ФНО-α, ИЛ-12 и ИЛ-18. ИНФ-γ обладает противовирусным действием, но для индукции этих эффектов является более слабым медиатором, чем ННФ типа I. Основной физиологической ролью ИНФ-γ является его способность модулировать иммунные реакции. ИНФ-γ индуцирует синтез многих белков, которые играют важную роль в презентации антигенов Т-клеткам, включая гликопротеины классов I и II ГКГ, инвариантную цепь, компоненты протеасомы Lmp2 и Lmp7 и внутриклеточные транспортеры пептидов ТАР1 и ТАР2.
Эти изменения усиливают эффективность презентации антигена Т-клетками CD4 и CD8. ИНФ-γ необходим также для активации макрофагов до их полного антимикробного потенциала, что помогает им уничтожать микроорганизмы, способные к внутриклеточному росту. Как и интерфероны типа I, ИНФ-γ обладает также сильным антипролиферативным действием на некоторые типы клеток. И, наконец, ИНФ-γ является индуктором избранных цитокинов (лигандов 9-11 хемокинов семейства СХС) и индуктором молекул адгезии эндотелиальных клеток (например, ICAM-1 и VCAM-1). Благодаря широкому спектру действий, ИНФ-γ является наиболее близким к первичному цитокину из всех остальных Т-клеточных цитокинов.
- Рекомендуем далее ознакомиться со статьей "Противовоспалительный цитокин интерлейкин-10 (ИЛ-10)"
Оглавление темы "Иммунология кожи.":- Цитокины интерлейкин-7 (ИЛ-7) и тимический стромальный лимфопоэтин
- Цитокины с рецепторами использующими бета-цепь интерлейкина-3 (ИЛ-3)
- Цитокины с рецепторами использующими гликопротеин 130: ИЛ-6, ИЛ-11, ИЛ-24, фактор ингибиции лейкоза, онкостатин М, цилиарный нейротропный фактор и кардиотропин-1
- Цитокины ИЛ-12, ИЛ-23, ИЛ-27, ИЛ-35
- Интерфероны как прототипы сигнализации через JAK/STAT
- Противовоспалительный цитокин интерлейкин-10 (ИЛ-10)
- Цитокины ИЛ-19, ИЛ-20, ИЛ-24, ИЛ-26
- Трансформирующий фактор роста β (TGF-β) и его рецептор
- Хемокины и терапевтическое применение цитокинов
- Хемокины кожи и их открытие