
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Фотостарение кожи - признаки, механизмы (патогенез)
Клинические и гистологические особенности поврежденной солнцем кожи перечислены в таблице ниже. Ведущей чертой кожи при фотостарении является эластоз, процесс, который характеризуется изменением цвета на желтый (диспигментация), шероховатостью и спутанными массами деградировавших эластических волокон, которые в дальнейшем разрушаются, формируя скопления, состоящие из дезорганизованного тропоэластина и фибриллина. Хотя фибриллин в изобилии содержится в эластическом веществе в глубине дермы, в ее верхних частях в области дермально-эпидермального соединения количество фибриллина снижается.
Кроме того, в фотоповрежденной коже увеличивается количество основного вещества, состоящего главным образом из GAG и протеиногликанов, тогда как количество коллагена снижается, отчасти из-за возрастающей активности металлопротеиназ. В отличие от защищенной от солнца стареющей кожи, в которой отмечается снижение общего числа клеток, подверженная фотостарению кожа часто характеризуется увеличением количества гиперпластических фибробластов и клеток воспаления, включая мастоциты, гистиоциты и другие мононуклеарные клетки, что стало причиной возникновения термина «гелиодерматит» (буквально, «воспаление кожи в результате воздействия солнца»). Иммуногистологические исследования выявили увеличение в дерме количества CD4+ Т-клеток.
В сосудистом русле кожи, подвергшейся фотостарению легкой степени, обнаруживается утолщение стенок венул, а при тяжелом фотоповреждении — утолщенные стенки сосудов содержащие поврежденные периваскулярные вуалевидные клетки (формирующиеся телеангиоэктазии).
В отличие от кожи при естественном старении, при фотостарении в эпидермисе часто обнаруживаются признаки акантолиза, хотя кроме потери полярности и клеточной атипии может отмечаться тяжелая атрофия. Также, наблюдается более выраженное уменьшение числа и снижение функции клеток Лангерганса. Дополнительные изменения приведены в таблице ниже.
Относительная тяжесть индуцированных солнцем повреждений кожи значительно варьирует у различных людей, отражая наследственные различия в уязвимости и способности к восстановлению после солнечного воздействия. Фотостарение отмечается не только у светловолосых лиц (кожные фототипы I и II), но и у лиц с более темными кожными фенотипами III и IV с обильной солнечной экспозицией в анамнезе. Обычно наиболее выражен процесс на лице, шее или разгибательных поверхностях верхних конечностей. Примечательно, что внешний вид фотоповрежденной кожи при I и II фототипах кожи отличается от более темных типов. У первых обычно отмечаются атрофические и диспластические изменения кожи с формированием актинического кератоза и злокачественных новообразований эпидермиса, в то время как у последних отмечается гипертрофический ответ: образование борозд, лентигиноза и шероховатости.
В одном исследовании было показано, что у пациентов с базальноклеточным раком отмечается меньшее количество морщин и тяжесть фотоповреждения по сравнению с лицами сходной комплекции. Это позволяет предположить, что определенные фенотипы фотостарения коррелируют с предрасположенностью к мутациям и канцерогенезу.
Фотостарение происходит не только у белокожих людей, но также у азиатов, латиноамериканцев и африканцев. Различия клинических проявлений при фотостарении у белокожих и других этнических групп прежде всего связаны с различиями защитных систем от УФ. В последних трех группах меланин является основной формой защиты, в то время как у белокожих меланин играет меньшую роль, а более значимым является утолщение рогового слоя. В одном из исследований было показано, что фактор защиты от солнечных лучей для эпидермиса темнокожих составляет 13,4 по сравнению с 3,4 для эпидермиса белокожих. Эпидермис чернокожих пропускает в дерму примерно 6% ультрафиолета В (УФВ), по сравнению с почти 30% в случае белой кожи. Сходным образом только приблизительно 18% УФА переносится в дерму темнокожих лиц по сравнению с более чем 55% у светлокожих.
Основными клиническими особенностями фотостарения кожи азиатов являются солнечное лентиго и пятнистая пигментация. Морщины от умеренной до тяжелой степени тяжести развиваются у азиатов только на шестом десятилетии жизни и только у лиц, регулярно проводивших много времени на солнце.
К настоящему времени не проводилось специальных исследований, посвященных фотостарению у латиноамериканцев, а результаты исследования по фотостарению темной кожи были опубликованы только в отношении афроамериканцев. Как и ожидалось, у более светлокожих латиноамериканцев и африканцев клинические признаки фотостарения возникали раньше и были более отчетливы, чем у лиц той же этнической группы с очень темной кожей. Изменения включают образование мелких морщин и пятнистую пигментацию.
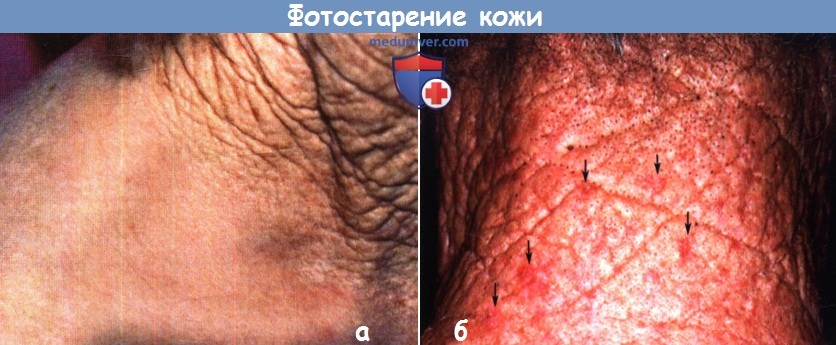
Подверженная частому воздействию солнечных лучей кожа выше воротниковой линии покрыта отчетливыми морщинами и обвислая,
что контрастирует с кожей нижней части шеи и плеча того же возраста, но защищенной от солнечных лучей.
Несмотря на разительные различия во внешнем виде, в обеих областях отмечаются возрастные функциональные изменения.
б - Фотостарение: гелиодерматит.
Выраженное образование борозд, желтая диспигментация и шероховатая поверхность (солнечный эластоз) с закупоренными фолликулами на шее.
Стрелками указаны небольшие участки актинического кератоза.
а) Механизмы фотостарения кожи. Механизмы, задействованные в естественном старении, также играют роль при фотостарении. Дополнительные механизмы обсуждаются ниже.
1. Передача сигнала через цитоплазматическую и ядерную мембрану. УФ-лучи посредством продукции активных форм кислорода, ингибирующих фосфатазы, поддерживающие рецепторы в неактивном состоянии активируют (фосфорилируют) рецепторы поверхности клеток, включая рецепторы эпидермального фактора роста, ИЛ-1 и фактор некроза опухолей (ФНО-а), что индуцирует передачу внутриклеточного сигнала, конечным результатом которого является активация ядерного транскрипционного комплекса АР-1, состоящего из белков c-Jun и c-Fos. В интактной коже человека даже субэритемогенные дозы УФВ [приблизительно 0,1 минимальной эритематозной дозы (МЭД)] посредством транскрипции регулируют и активируют АР-1.
Повышение активности АР-1 снижает синтез главных коллагенов дермы I и III типа посредством блокировки действия трансформирующего фактора роста (TGF-α) — цитокина, усиливающего транскрипцию гена коллагена. АР-1 также снижает уровень рецепторов TGF-α, что еще в большей степени ингибирует транскрипцию коллагена и антагонистически действует на эффекты естественных ретиноидов в коже, приводя к развитию функционального дефицита ретиноидов и снижению синтеза коллагена, в норме стимулируемого ретиноевой кислотой, связывающейся с ее ядерными рецепторами. Кроме того, УФ провоцирует синтез и секрецию богатого цистеином регуляторного фактора роста (CYR61), который снижает синтез проколлагена I типа, повышает уровни ММР-1, снижает уровень рецептора TGF-β и вызывает активацию АР-1. Таким образом, в поврежденной УФ излучением коже отмечается общая редукция синтеза коллагена. Повышенная активность АР-1 также повышает уровни и активность нескольких ферментов, разрушающих компоненты внеклеточного матрикса, особенно ММР-1 (коллагеназа), ММР-3 (стромелизин-1) и ММР-9 (92-kd желатиназа). УФ также активирует нуклеарный фактор кВ, фактор транскрипции, провоцирующий экспрессию множества белков, о которых упоминалось в разделе «Механизмы естественного старения кожи», и усиливает разрушение дермального матрикса за счет возрастающих уровней ММР-1 и ММР-9.
Далее разрушение матрикса усиливается ММР-8 (коллагеназа) нейтрофильного происхождения, что происходит в результате нейтрофильной инфильтрации облученных УФ зон. Хотя также отмечается сопутствующая активация тканевых ингибиторов металлопротеиназ (TIMP), ограничивающих разрушение матрикса, TIMP видимо не в состоянии полностью блокировать суммарное повреждение коллагена кожи. УФ уменьшает также уровень FoxO mRNA, еще более нарушая синтез коллагена I и повышая транскрипцию ММР-1 и -2 (18С).
Индуцированная УФ деградация коллагена обычно не полная и приводит к накоплению в дерме частично разрушенных фрагментов коллагена, вызывая таким образом снижение структурной целостности кожи. Кроме того крупные продукты распада коллагена ингибируют синтез нового коллагена и, таким образом, само по себе разрушение коллагена негативным образом сказывается на синтезе нового коллагена. Примечательно, что при естественном старении отмечаются связанная со стрессом повышенная активность АР-1, повышение уровней CYR61 и ММР и снижение продукции коллагена. Это позволяет предположить, что сходные механизмы вносят вклад и в хронологическое старение, возможно, опять же в результате образования активных форм кислорода, как это обсуждалось в отдельной статье на сайте «Механизмы естественного старения кожи».
УФ-излучение, как прямо, так и посредством генерирования реактивных форм кислорода (РФК) индуцирует транскрипцию тропоэластина, компонента зрелых эластических волокон. В эластическом материале повышено также содержание фибулинов-2 и -5 и фибриллина-1, компонентов микрофибриллярной фракции эластических волокон дермы. Кроме того, в фотоповрежденной коже отмечаются повышенные уровни эластазы в результате синтеза и секреции эластазы нейтрофилами, привлеченными в участок повреждения медиаторами воспаления. Таким образом, избыточный, разбалансированный синтез компонентов эластических волокон, подвергающихся частичной деградации, приводит к образованию аморфного эластотического материала.
2. Митохондриальное повреждение. Митохондрии представляют собой органеллы клеток, вырабатывающие энергию при поглощении кислорода (аденозин трифосфат). Несмотря на то что они имеют антиоксидантные защитные системы, постоянно образующиеся активные формы кислорода повреждают митохондриальную ДНК (мтДНК). На сегодняшний день механизмы репарации поврежденной ДНК в митохондриях не обнаружены, хотя у них наблюдается способность к базовому восстановлению повреждений и восстановлению оксидативных повреждений. Тем не менее частота мутаций мтДНК приблизительно в 50 раз выше, чем в ядерной ДНК, а кожа при фотостарении имеет большее число мутаций мтДНК по сравнению с защищенной от солнца кожей, обнаруживая крупные делеции ДНК. Это приводит к снижению функции митохондрий, что в дальнейшем вызывает накопление активных форм кислорода и нарушает способность клеток вырабатывать энергию.
Также была обнаружена корреляция между сниженной функцией митохондрий и повышением уровня ММР-1 без сопутствующего повышения ММР-1-специфичного TIMP, что вызывало деградацию коллагена и усиливало фотостарение кожи.
3. Окисление белков. Белки подвергаются оксидативному повреждению, и при фотостарении в верхних частях дермы отмечается накопление окисленных поврежденных белков. В исследованиях in vitro предполагается роль УФА как главного триггера появления и накопления таких белков, что в дальнейшем ингибирует протосомальную функцию и способность клеток к успешному разрушению вновь поврежденных белков.
4. Повреждение базальной мембраны. В открытой воздействию солнца коже базальная мембрана утолщается и становится многослойной частично в результате повреждения, вызванного активацией матриксных металлопротеиназ, которые воздействуют на молекулярный транспорт между эпидермисом и дермой и ухудшают состояние эпидермиса.
б) Спектр действия при фотостарении. Спектр действия при фотостарении у человека не определен, однако многие исследователи изучали относительный вклад различной длинны волн светового спектра на моделях животных.
Несмотря на четко зафиксированные эффекты УФВ, предполагается, что роль УФА в фотостарении кожи пропорционально более значительная, поскольку, по сравнению с УФВ, наличие УФА в земном солнечном свете как минимум в 10 раз больше, объем годового излучения и среднего дневного излучения также намного выше, больше также средняя глубина проникновения УФА в дерму.
В коже грызунов эластозо-подобные состояния могут быть вызваны длительным интенсивным воздействием преимущественно УФА или УФВ источника. Фотоны УФВ в среднем в 1000 раз более активны, чем фотоны УФА, и прежде всего ответственны за развитие солнечных ожогов, загара и фотоканцерогенез в результате пребывания на солнце. УФВ являются главной причиной прямого повреждения ДНК и вызывают воспаление и иммуносупрессию, а также синтез и высвобождение постагландинов (PG), особенно PGE2, посредством индукции фермента циклооксигеназы-2. УФВ также индуцирует орнитин декарбоксилазу, фермент, лимитирующий биосинтез полиаминов, которые стимулируют пролиферацию клеток (участвующих в развитии рака) и вносят вклад в кожный ангиогенез посредством снижения экспрессии ингибитора ангиогенеза тромбоспондина-1 и индукции экспрессии сосудистого эндотелиального фактора роста и выделенного из тромбоцитов фактора роста эндотелиальных клеток (двухангиогенных факторов). УФВ также усиливает проникновение в кожу продуцирующих эластазу лейкоцитов, ускоряя деградацию эластина.
Более того, кожа человека, ежедневно в течение месяца облучаемая субэритематозными дозами УФА, характеризуется гиперплазией эпидермиса, утолщением рогового слоя, уменьшением количества клеток Лангерганса и формированием воспалительных инфильтратов в дерме с отложением на эластических волокнах лизоцима. УФА-излучение также индуцирует синтез и высвобождение цитокинов и ММР, особенно коллагеназы (ММР-1) и эластазы, а также вызывает мутации митохондриальной ДНК. Как УФА, так и УФВ приводят к образованию активных форм кислорода, повреждающих клеточные липиды, белки и ДНК. Исследования с использованием лазерной микродиссекции человеческой кожи показали, что кератиноциты базального слоя с мутантным геном р53 в дополнение к характерным для УФВ-мутациям отличаются большим числом мутаций, связанных с УФА (и главным образом фотоповреждения 8-OXO-dG). Это позволяет предполагать, что УФА является важным этиологическим фактором в развитии базальноклеточного рака. УФА-излучение более эффективно в индукции окислительных повреждений, чем УФВ. Действительно, степень солнечного эластоза и кожного фотостарения, по-видимому, коррелируют с уровнем накопительного окисления белков дермы, но не окисления в эпидермисе. Предполагается, что это указывает на более развитую антиоксидантную сеть и/или лучшую восстановительную способность эпидермиса.
Солнечный свет также содержит инфракрасное излучение (ИК) (от 760 нм до 1 мм). Излучение с длиной волны от 760 до 1400 нм может проникать в кожу и достигать подкожных тканей, не вызывая выраженного повышения температуры кожи. Напротив, излучение с длиной волны от 1400 нм до 1 мм в основном поглощается эпидермисом и значительно повышает температуру кожи. ИК излучение имеет значение главным образом в областях с высокой теплоизоляцией и усиливает вызванные УФА изменения в дерме, приводя к развитию эластоза тяжелой степени. Даже при воздействии без УФ ИК излучение влияет на эластические волокна дермы и повышает количество основного вещества дермы. На моделях безволосых мышей было показано, что ИК при совместном облучении вносит вклад в вызванное УФВ утолщение эпидермиса и дермы, а в отдельности от УФ — индуцирует экспрессию ММР-3 и мышиного эквивалента ММР-1. Более того, в коже человека в результате воздействия I IК увеличивается экспрессия тропоэластина, главного компонента эластических волокон, который соединяется с микрофибриллами, а ИК индуцирует экспрессию фибриллипа-1, компонента микрофибрилл.
Кроме того, повышается уровень ММР-12, фермента, расщепляющего эластин. Таким образом, волны ИК спектра, по-видимому, вносят вклад в вызванное УФ излучением фотостарение.
Многие связанные с возрастом физиологические изменения, такие как замедленное заживление ран и снижение иммунореактивности, также, по-видимому, ускоряются в коже, подверженной солнечному воздействию. Более того, клетки, выделенные из областей кожи, длительное время подверженных воздействию солнечных лучей, отличаются от клеток, выделенных из защищенных от солнца участков кожи тех же доноров, за счет укороченного времени жизни культуры, замедления роста, более низкой плотности популяции и измененной реактивностью в ответ на воздействие ретиноевой кислоты. Все изменения находились в зависимости от хронологического возраста донора. Предполагается, что несколько известных механизмов, принимающих участие в вызванном УФ повреждении клеток, также лежат в основе хронологического старения, хотя соответствующие изменения отмечаются только после седьмого десятилетия жизни. Они включают повреждение ДНК и/или ухудшение ее репарации, окислительное повреждение, лизосомальное разрушение, повышение уровня ММР, снижение продукции коллагена и повреждение соединительной ткани.

- Вернуться в оглавление раздела "дерматология."
Оглавление темы "Старение кожи.":- Причины и механизмы старения кожи (патогенез)
- Естественное старение кожи - причины, механизмы, гистология (патогенез, морфология)
- Естественное старение волос - причины, механизмы (патогенез)
- Фотостарение кожи - признаки, механизмы (патогенез)
- Связь болезней кожи с возрастом человека
- Опухоли кожи пожилых - типы, признаки
- Заболевания кожи пожилых - типы, признаки