
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Культивирование эмбриональных стволовых клеток. Участие стволовых клеток в эмбриогенезе
При крайне ограниченных фенотипических признаках и отсутствии большинства блоков транссигнализации ЭСК все же экспрессируют некоторые рецепторные молекулы, которые можно использовать для их идентификации. Примечательно, что антигены-маркеры ЭСК у человека и приматов оказались общими. Чаше всего для маркировки ЭСК используют меченые антитела к мембранносвязанным антигенам SSEA-3, SSEA-4 (уникальные липидные антигены, представляющие комплекс гликолипида GL7 с сиаловой кислотой), а также высокополимерные гликопротеиды TRA-1-81, TRA-1-60 (Репин, 2001).
Кроме того, стволовые клетки экспрессируют специфический эмбриональный антиген SSEA-1 и эндогенную щелочную фосфатазу (Resnick et al., 1992), а также специфический транскрипционный фактор Oct4 (Scholer et al., 1990). Последний необходим для поддержания механизмов пролиферации ЭСК — специфический транскрипционный фактор Oct4 активирует экспрессию гена фактора роста фибробластов 4 и стабилизирует экспрессию бокса генов, ответственных за нелимитированную редупликацию ДНК в незрелых клетках (Nichols et al., 1998).
Наиболее важными внутриклеточными маркерными белками являются Oct3, Oct4, Tcf и Groucho, относящиеся к белкам-сайленсерам хроматина (Репин, 2001).
Практически сразу после того как многолетние попытки культивировать стволовые клетки вне организма увенчались успехом и были получены первые культуры стволовых клеток, выделенных из бластоцист мыши (Evans, Kaufman, 1981; Martin, 1981), а также культуры первичных половых клеток (Donovan, 1994), начался этап исследований плюрипотентного потенциала ЭСК при их введении в эмбрионы на ранних стадиях развития. Было показано, что на стадии морулы и бластоцисты ЭСК способны формировать химерные зародыши, в которых потомки донорских ЭСК выявляются во всех соматических тканях и даже в гаметах (Robertson, Bradley, 1986; Allan, 1987; Robertson, 1987; Joyner, 1993; Hoganet al, 1994). Таким образом, в биологии развития с помощью ЭСК был установлен "мост" между экспериментальными исследованиями in vivo и in vitro, что значительно расширило возможности изучения процессов закладки первичных тканей и органов, их дифференцировки и эмбрионального органогенеза (Серов и др., 2001).
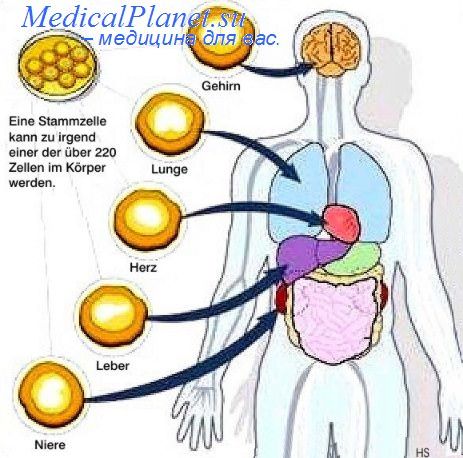
Четко установлено, что in vivo в процессе эмбриогенеза ЭСК интегрируются в клеточную массу раннего зародыша, а их производные обнаруживаются во всех органах и тканях. ЭСК колонизируют в химерном зародыше линию половых клеток, потомки которых образуют полноценные яйцеклетки и спермин. Эмбриональные стволовые клетки клоногенны — единичная ЭСК способна создать генетически идентичную колонию клеток с молекулярными маркерами, к которым относится экспрессия гена oct4 и щелочной фосфатазы, высокая активность теломеразы, а также экспрессия определенных эмбриональных антигенов.
Для изучения механизмов эмбриогенеза с помощью стволовых клеток разработана методика химеризации морулы путем создания биологической конструкции, снаружи которой располагается слой тетраплоидных бластомеров реципиента, а внутрь вводятся донорские ЭСК. Таким образом, трофобласт формируется из потомков тетраплоидных бластомеров реципиента, что обеспечивает имплантацию и плацентацию, а донорские ЭСК выступают в роли внутренней клеточной массы, из которой образуется тело жизнеспособного зародыша и линия первичных прогениторных половых клеток (Дыбан А., Дыбан 77., 2002).
Исследовательская ценность эмбриональных стволовых клеток заключается не только в том, что при манипуляциях in vitro с их геномом сохраняется плюрипотентность, но и в том, что при этом сберегается способность ЭСК участвовать в образовании первичных половых клеток химерного зародыша. Показано, что потомки всего одной генетически модифицированной ЭСК заселяют все первичные зачатки и формирующиеся ткани химерного эмбриона, полученного посредством агрегации или кокультивирования этой клетки с 8-клеточным зародышем (Saburi et. al., 1997). При трансплантации в морулу мышей ЭСК, трансфецированных геном зеленого флуоресцентного протеина, флуоресцирующие потомки этой клетки были обнаружены во всех исследуемых тканях развивающегося эмбриона (Shimada, 1999). Трансплантация ЭСК в морулу позволяет создавать жизнеспособных мышей, организм которых состоит только из потомков донорской ЭСК (Nagy, 1990; Eggan, 2001), что открывает перспективы для различных вариантов терапевтического клонирования.
Сейчас такой методический подход успешно применяется для изучения проблем биологии развития, в частности, с его помощью проводят анализ механизмов генетической инактивации Х-хромосомы или эпигенетической нестабильности ЭСК. Трансплантация ЭСК в ранний зародыш используется и в биотехнологиях сельского хозяйства, а также в экспериментах по генотерапии (Дыбан А., Дыбан П., 2002).
- Читать далее "Генетически модифицированные стволовые клетки. Участие генов в дифференциации стволовых клеток"
Оглавление темы "Дифференциация эмбриональных стволовых клеток":1. Культивирование эмбриональных стволовых клеток. Участие стволовых клеток в эмбриогенезе
2. Генетически модифицированные стволовые клетки. Участие генов в дифференциации стволовых клеток
3. Гибридные клетки. Цитогибриды
4. Видовые эмбриональные гибридные клетки. Получение клонов гибридных клеток
5. Хромосомный и генетический набор гибридных клеток. Плюрипотентность гибридных клеток
6. Репрограммирование Х-хромосомы гибридных клеток. Плюрипотентность цитогибридов
7. Хромосомная память. Эффективная экспансия стволовых клеток в культуре
8. Спонтанная дифференцировка эмбриональных стволовых клеток. Дифференцировка стволовых клеток
9. Индуцированная дифференциация стволовых клеток. Направленная дифференциация стволовых клеток
10. Селективная дифференциация стволовых клеток. Регуляция дифференцировки стволовых клеток