
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Дополнительная моторная кора. Функции дополнительной моторной коры.
Термин «дополнительная моторная кора» (ДМК) был впервые введен в 1949 г. W.Penfield и K.Welch для обозначения части поля 6 (по Brodmann) на медиальной поверхности полушарий, стимуляция которой вызывала сложные движения и позы. ДМК получает массивную афферентацию от ассоциативных зон префронтальной и теменной коры, а также от первичной и вторичной соматосенсорных зон и ПМК.
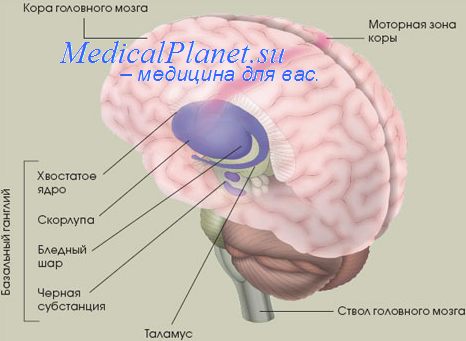
Кроме того, через таламус в дополнительной моторной коре стекается основная часть афферентации базальных ганглиев, в меньшей степени — афферентация от мозжечка (главным образом, от контралатерального зубчатого ядра).
Эфферентные пути от дополнительной моторной коры направляются в основном в ипсилатеральную ПМК, базальные ганглии и мозжечок, а через мозолистое тело — в контралатеральные моторные зоны коры. Только 5% нейронов ДМК связаны прямыми моносинаптическими путями с мотонейронами передних рогов спинного мозга. Эти нейроны регулируют движения в дистальных отделах рук (независимо от ПМК) и при поражениях ПМК могут частично брать на себя ее функцию.
Основная функция дополнительной моторной коры заключается в подготовке движений — трансформации намерения в конкретную последовательность моторных профамм. Об этом свидетельствует тот факт, что многие нейроны ДМК селективно разряжаются в момент получения инструкции, но прекращают активность с началом движения. Причем эта активность, в отличие от активности нейронов ПМК, мало коррелирует с параметрами предстоящего движения (например, с силой сокращения мышц).
Регистрация ЭЭГ в период подготовки к произвольному движению (за 0,5—1 с до начала мышечного сокращения) выявляет симметричный медленный негативный потенциал (потенциал готовности — Bereis-chaftpotential), который имеет наибольшую амплитуду над областью верхушки. В прецентральных областях потенциал готовности симметричен только в первой фазе, а затем становится асимметричным за счет большей выраженности в контралатеральном по отношению к работающей конечности полушарии.
Потенциал готовности, по-видимому, отражает тоническую активацию дополнительной моторной коры (с обеих сторон — даже при одностороннем движении!) и, возможно, ПреМК, а также префронтальной и передней поясной коры, в меньшей степени ПМК (с контралатеральной стороны). ПМК ифает важную роль в генерации более поздних латерализо-ванных компонентов потенциала готовности. Потенциал готовности регистрируется только перед спонтанными движениями и отсутствует перед пассивными или рефлекторными (выполняемыми в ответ на появление определенного сенсорного стимула) движениями.
С началом движения «подготовительная» тоническая активность дополнительной моторной коры прекращается. Этот феномен можно объяснить тем, что начатое движение способно выступать как триггер, инициирующий вспышку активности в базальных ганглиях, которая в конечном итоге «докатывается» до БШв и ЧСр и приводит к торможению соответствующих таламокорктикальных нейронов, проецирующихся на ДМК. Прекращение тонической активности ДМК не только позволяет реализоваться начатому движению, но и создает условия для начала подготовки нового этапа движения или следующего движения.
Таким образом, взаимодействие базальных ганглиев и дополнительная моторная кора обеспечивает переключение с одной моторной профаммы на другую.
Данные электрофизиологических исследований и функциональной нейровизуализации показывают, что ДМК совместно с ПМК, базальными ганглиями (задними отделами скорлупы) и мозжечком преимущественно вовлечены в выполнение заученных автоматизированных или последовательных многоэтапных движений, имеющих сложную временную организацию и выполняемых спонтанно, по памяти, без пусковых стимулов и «опоры» на внешние ориентиры. То есть тех движений, которые нуждаются в тщательном внутреннем планировании и более сложной подготовке.
В этом основное отличие дополнительной моторной коры от ПреМК, которая (совместно с префронтальной корой, задними отделами теменной коры и передней частью стриатума) играет ведущую роль при выполнении новых, необычных для данного человека движений, выполнение которых требует внимания, интефации сенсорной информации различной модальности и функционирования оперативной памяти (Alexander G.A., DeLong M.R., 1992).
Нейроны дополнительной моторной коры (но не ПреМК) селективно разряжаются во время выполнения или перед выполнением определенной последовательности движений, а методы функциональной нейровизуализации выявляют повышение активности в ДМК при заучивании многоэтапных движений. В клинике при поражениях ДМК страдает не только выполнение уже заученных движений, но и приобретение новых двигательных навыков.
Имея двусторонние проекции, дополнительная моторная кора играет важную роль в координации движений, вовлекающих обе стороны тела. При стимуляции ДМК чаще возникают сложные медленные двусторонние движения, но возможны и односторонние движения. При регистрации активности отдельных нейронов показано, что некоторые клетки ДМК преимущественно связаны с контралатеральными сенсомоторными зонами, но часть нейронов разряжается только при координированных двусторонних движениях пальцев, но не при движении одной рукой.
Дополнительная моторная кора, как и дорсальная часть ПреМК, взаимодействуя с задней теменной корой, может инициировать изменение позы, которое предшествует движению конечности и имеет цель удержать равновесие (если планируемое движение ему угрожает). Важное значение ДМК имеет и в продукции речи, что подтверждается результатами очаговых поражений ДМК доминантного полушария, вызывающими мутизм, а также данными функциональной нейровизуализации. При стимуляции этой области могут возникать вокализации или остановка речи.
Исследования на приматах показали, что дополнительная моторная кора может модулировать длиннолатентные транскортикальные рефлексы, тормозя реакции нейронов ПМК на соматосенсорную афферентацию. Высвобождением реакций на соматосенсорные стимулы от этого ингибиторного контроля объясняется появление при поражении ДМК хватательного рефлекса. При поражении ДМК возникают неадекватные позные установки, неловкость при перемещении конечностей, нарушения программирования длиннолатентных рефлексов, расстройство бимануальной координации. Дисфункция ДМК в результате первичного повреждения или разобщения с базальными ганглиями может лежать в основе акинезии (гипокинезии), характерной для паркинсонизма.
В настоящее время в пределах 6-го поля выделяют две зоны: ростральную (ПредДМК) и каудальную (собственно ДМК). Хотя ПредДМК активируется в момент подготовки и выполнения движений, она не имеет прямых связей с ПМК или спинным мозгом, а также соматотопической организации, и поэтому ее формально нельзя отнести к моторным зонам коры. Но тесные реципрокные связи ДМК и предДМК указывают, что последняя может играть роль в высшем моторном контроле. ПредДМК также имеет тесные связи с префронтальной и передней поясной корой.
В ПреДМК поступает афферентация и от мозжечка, но не от базальных ганглиев. Нейроны предДМК (как и нейроны собственно ДМК) чаще реагируют на зрительные, чем на соматосенсорные стимулы.
Стимуляция предДМК может вызвать императивный позыв к выполнению движения (который отсутствует при стимуляции ПМК) или ощущение неминуемого движения. Активация предДМК происходит даже при мысленном представлении движения. ПреДМК может быть той зоной, где происходит согласование внешних стимулов или команд с внутренними потребностями и желаниями индивида, реализующееся в определении стратегии и планировании произвольных действий. ПредДМК может играть важную роль в заучивании новой последовательности движений.
Однако по мере закрепления и автоматизации двигательного навыка активность ПредДМК снижается, и при последующей тренировке контроль за движением постепенно перемещается от ДМК к ПМК.
- Вернуться в оглавление раздела "неврология"
Оглавление темы "Стриатум. Кора больших полушарий.":1. Нейроны стриатума. Матрикс стриатума.
2. Вентральный стриатум. Стриатопаллидарные связи. Субталамическое ядро.
3. Внутренний сегмент бледного шара. Таламические проекции базальных ганглиев.
4. Связи таламуса со стриатумом. Как функционируют стриато-паллидо-таламо-кортикальные связи?
5. Компактная часть черной субстанции. Нигростриарная система.
6. Физиология дофамина. Влияние дофамина на нигростриарную систему.
7. Стволовые проекции базальных ганглиев. Связи базальных ганглиев с двигательными ядрами.
8. Роль коры больших полушарий в регуляции движений. Первичная моторная кора.
9. Премоторная кора. Строение прематорной коры. Признаки поражения прематорной коры.
10. Дополнительная моторная кора. Функции дополнительной моторной коры.