
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Нейроны стриатума. Матриркс стриатума.
Стриатум содержит также не менее четырех типов вставочных нейронов: 1) крупные холинергические нейроны (составляют 1—2% от клеточной популяции стриатума), 2) ГАМКергические нейроны, 3) нейроны, содержащие соматостатин, нейропептид Y и синтетазу оксида азота, 4) нейроны, содержащие кальретинин (Бархатова В.П., 1988; Леонтович Т.А., 1984; Wichmann Т., DeLong M.R., 1997).
Вставочные нейроны получают корковую и таламическую афферентацию и разряжаются спонтанно, вне связи с подготовкой или выполнением определенных движений, являясь источником тонической активности стриатума. Многочисленные коллатерали их аксонов контактируют с отростками или телами проекционных шиловидных нейронов и обычно тормозят их активность (Раева СН, 1977; Alexander G. Е., 1997).
Нейроны стриатума образуют скопления (стриосомы), занимающие примерно 15% площади стриатума и окруженные зонами с менее плотным расположением клеток (матрикс). Для стриосом характерны низкая активность ацетилхолинэстеразы, высокое содержание дофамина, динорфина, нейротензина, мю-опиатных рецепторов, преобладание D1-рецепторов.
Матрикс отличается высокой активностью ацетилхолинэстеразы, высоким содержанием соматостатина и энкефалина, преобладанием D2-рецепторов (Afifi А.К., Bergman R.A., 1998). Стриосомы сконцентрированы в медиовентральной части стриатума, матрикс наиболее гомогенен в его дорсолатеральной части. Стриосомные ГАМКергические нейроны, созревающие в процессе индивидуального развития раньше, чем матриксные нейроны, получают афферентацию преимущественно от лимбических структур (прежде всего от миндалины), а также от передней поясной коры, задней орбитофронтальной, медиофронтальной и других ассоциативных зон коры.
Проецируются они главным образом на нигростриарные нейроны ЧСк, аксоны которых в свою очередь подходят к нейронам как стриосом, так и матрикса.
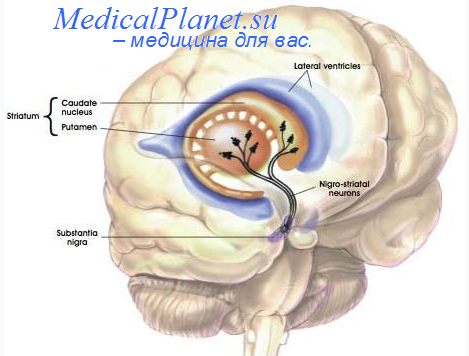
Нейроны матрикса в различных зонах стриатума получают афферентацию главным образом от моторных и сенсорных зон коры, а также ассоциативной префронтальной коры и интраламинарных ядер таламуса и направляют аксоны к БШн, БШв, ЧСр и СТЯ. В пределах матрикса выделяют скопления клеток, получающих общую афферентацию (матрисомы).
При этом ограниченные зоны коры проецируются на различные стриосомы или матрисомы, и наоборот, каждая стриосома или матрисома связана с несколькими корковыми зонами (Alexander G.E., 1997). Предполагают, что нейроны стриосом участвуют в оценке внешних и внутренних сигналов с точки зрения текущих потребностей индивидуума, тогда как нейроны матрикса задействованы в выборе адаптивных (в данных условиях) моторных программ.
Синхронизация активности нейронов матрикса и стриосом осуществляется холинергическими вставочными клетками, дендриты которых контактируют как с матриксными, так и со стриосомными нейронами, а аксоны подходят лишь к клеткам матрикса. Холинергические нейроны получают также глутамергическую афферентацию от интраламинарных ядер таламуса и дофаминергическую афферентацию от нейронов среднего мозга. Синхронизирующая активность холинергических вставочных нейронов нарастает в процессе обучения при условии положительного подкрепления (под влиянием дофаминергической афферентации).
Таким образом, на уровне стриатума может происходить интеграция афферентации от различных отделов коры, обеспечивающая, например, мотивационноаффективную поддержку тех или иных действий, планируемых в данной конкретной ситуации. В синхронизации активности стриарных проекционных нейронов участвуют и ГАМКергические вставочные нейроны, получающие афферентацию как от корковых, так и паллидарных нейронов и обеспечивающие таким образом как прямую, так и обратную связь (Alexander G.A., DeLong M.R., 1992).
В заключении следует подчеркнуть, что проекционные нейроны стриатума занимают стратегическое положение в системе базальных ганглиев — их активация зависит от взаимодействия импульсации из различных корковых и подкорковых источников и, в свою очередь, предопределяет изменение активности связанных с ними нейронов других базальных ганглиев.
Таким образом стриатум функционирует как гибко адаптирующийся (самонастраивающийся) фильтр, облегчающий или блокирующий на подкорковом уровне инициацию тех или иных корковых программ в зависимости не только от внутренних потребностей и внешней ситуации, но и от предшествующего опыта.
- Читать далее "Вентральный стриатум. Стриатопаллидарные связи. Субталамическое ядро."
Оглавление темы "Стриатум. Кора больших полушарий.":1. Нейроны стриатума. Матриркс стриатума.
2. Вентральный стриатум. Стриатопаллидарные связи. Субталамическое ядро.
3. Внутренний сегмент бледного шара. Таламические проекции базальных ганглиев.
4. Связи таламуса со стриатумом. Как функционируют стриато-паллидо-таламо-кортикальные связи?
5. Компактная часть черной субстанции. Нигростриарная система.
6. Физиология дофамина. Влияние дофамина на нигростриарную систему.
7. Стволовые проекции базальных ганглиев. Связи базальных ганглиев с двигательными ядрами.
8. Роль коры больших полушарий в регуляции движений. Первичная моторная кора.
9. Премоторная кора. Строение прематорной коры. Признаки поражения прематорной коры.
10. Дополнительная моторная кора. Функции дополнительной моторной коры.