
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Репродуктивные варианты вирусов. Условно-дефектные вирусы
Псевдовирусы. Псевдовирусы представлены вирусными капсидами, но одни из них - полые без вирусного генома, можно сказать вирусные тени, или вирусы-пустышки, а другие вместо вирусного генома содержат нуклеиновую кислоту клетки-хозяина. Образование неполных псевдовирионов объясняют перепроизводством вирионных белков, а псевдовирионов с нуклеиновой кислотой клетки - нарушением белокнуклеинового механизма «узнавания» при сборке вирусных частиц, т. е. их возникновение объясняют случайными обстоятельствами.
Вирусы-мутанты. Во время репликации вирусов спаривание правильного азотистого основания с основанием в таутомерной форме приводит к простой замене (транзиции) пурина на пурин или пиримидина на пиримидин. Скорость спонтанного мутагенеза в геномах ДНК-вирусов составляет 10~8 - 10-11 на каждый включенный нуклеотид, а в РНК-геномах - 10-3- 10-4.
Более высокие скорости накопления мутаций у РНК-содержащих вирусов, вероятно, связаны с меньшей корректирующей активностью РНК-полимераз, чем у ферментов, реплицирующих ДНК. Появляющиеся при этом мутанты, как правило, являются делеционными (лат. deletio - выпадение), т. е. утрачивающими определенный участок генома родительского вируса. Вирусные частицы с таким дефектным геномом сохраняют свою активность, но для репликации и созревания нуждаются в продуктах вирусного генома родителя - обычно в структурных и неструктурных белках.
Такой характер воспроизводства вирусов называют негенетическим типом взаимодействия или односторонней комплементацией (дополнением); родительский вирус, стимулирующий репродукцию мутанта, - вирусом-помощником, а репродуцирующийся с его помощью мутант - вирусом-сателлитом (спутником).
У ts-мутантов нуклеотидная последовательность в геноме изменяется таким образом, что образованный ими белковый продукт сохраняет функционально активную конформацию только при пермиссивной (англ. permissive - разрешающий) температуре около 36-38 °С, а при более высокой непермиссивной температуре 39-42 С мутант становится нежизнеспособным и прекращает развитие. Наоборот, tc-мутанты размножаются при более высокой, чем оптимальная, пермиссивной для них температуре.
В меньшей мере обоснован механизм мутации по спектру хозяев и морфологии бляшек, связанныйс какой-то особенностью их метаболизма. Мутации по спектру хозяев чаще идут в направлении его сужения, но некоторые мутанты приобретают способность к репродукции в не свойственных для дикого типа клетках.
Столь же разнообразны мутации по морфологии бляшек: мутанты аденовирусов с большими бляшками, например, быстрее, чем дикий тип, освобождаются из клетки, а синцитиальный мутант вируса простого герпеса вместо типичного цитопатическо-го действия приобретает способность индуцировать образование многоядерных клеток.
- Читать далее "Дефектные интерферирующие вирусы. Интеграционные вирусы с дефектным геномом"
Оглавление темы "Репродукция вирусов":1. Способы культивирования вирусов. Культуры клеток
2. Репродуктивные варианты вирусов. Условно-дефектные вирусы
3. Дефектные интерферирующие вирусы. Интеграционные вирусы с дефектным геномом
4. Рекомбинантные вирусы. Генная инженерия
5. Конструирование генов. Выделение генов
6. Синтез генов. Передача и клонирование генов
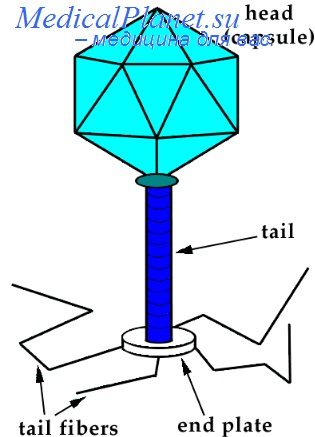
7. Классификация, форма и строение фагов. Свойства фагов
8. Взаимодействие вирулентного фага с бактерией. Умеренные фаги
9. Трансдукция фагов. Характеристика трансдуцирующих фагов
10. Абортивная трансдукция. Происхождение и распространение фагов