
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Строение, функции хроматина
Хроматин в неделящихся ядрах в действительности представляет собой хромосомы в различной степени раскручивания (деконденсации). В соответствии со степенью конденсации хромосом как под световым, так и под электронным микроскопом различают два типа хроматина. Гетерохроматин (греч. heteros — другой + chroma — цвет) обладает высокой электронной плотностью; под электронным микроскопом он имеет вид крупных гранул, а под световым микроскопом — базофильных глыбок.
Эухроматин соответствует менее скрученным участкам хромосом, которые под электронным микроскопом видны как мелкодисперсный зернистый материал, а под световым микроскопом — как светлоокрашенные базофильные участки. Пропорции гетерохроматина и эухроматина определяют соотношение темных и светлых участков ядер в срезах тканей при световой и электронной микроскопии. Интенсивность окрашивания хроматина в ядре часто учитывают как признак для разделения и идентификации отдельных тканей и типов клеток под световым микроскопом.
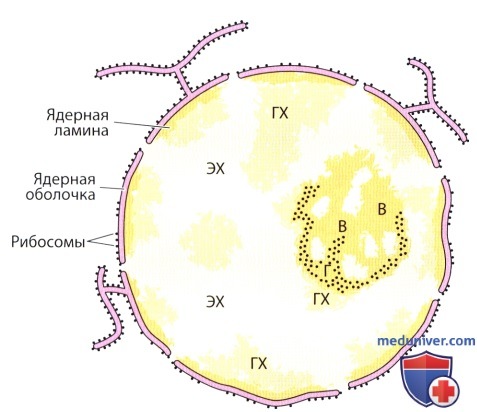
Ядерная оболочка состоит из двух мембран эндоплазматической сети, охватывающих перинуклеарную цистерну.
В местах слияния двух мембран образуются ядерные поры. К наружной ядерной мембране прикреплены рибосомы.
Глыбки гетерохроматина (ГХ) связаны с ядерной пластинкой, тогда как эухроматин (ЭХ) рассеян по внутренней части ядра.
В ядрышке обратите внимание на ассоциированный с ним хроматин, гетерохроматин (ГХ), гранулярную часть (Г) и волокнистую часть (В).
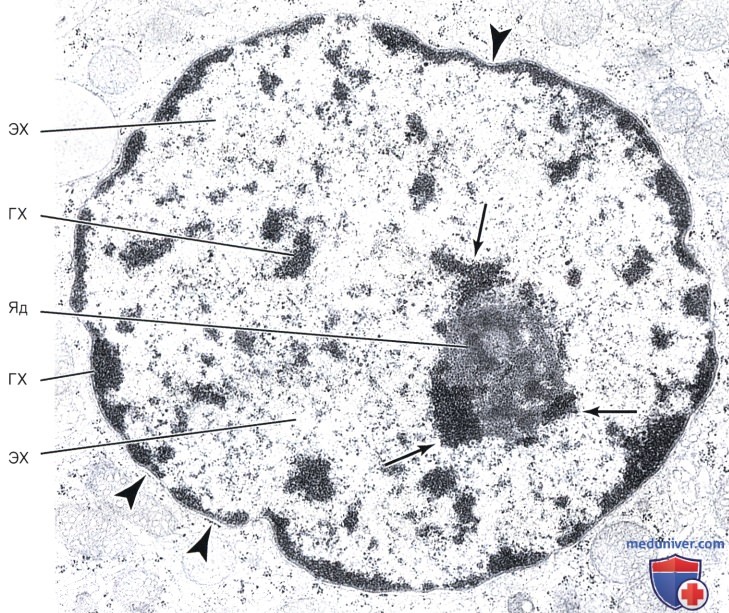
Стрелки без обозначений указывают на ассоциированный с ядрышком хроматин, расположенный вокруг ядрышка (Яд).
Треугольники указывают на перинуклеарную цистерну. Под цистерной располагается слой гетерохроматина, который является главным компонентом структуры, которая известна как ядерная мембрана и видна под световым микроскопом.
Электронная микрофотография, х26 000.
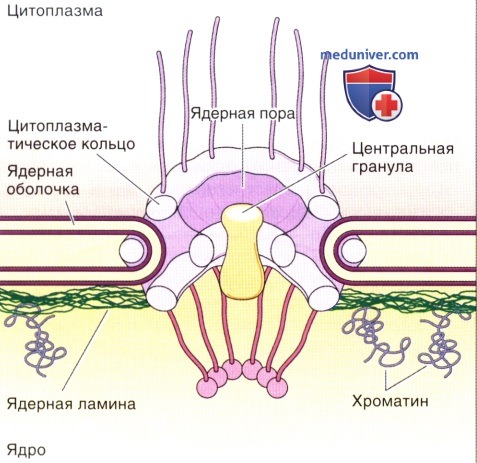
Комплекс ядерный поры состоит из двух белковых колец в виде восьмиугольных структур.
Из цитоплазматического кольца в цитозоль проникают длинные филаменты, а от внутриядерного кольца отходят филаменты, образующие корзинчатую структуру.
Присутствие центрально расположенной цилиндрической гранулы в ядерной поре признают не все исследователи.
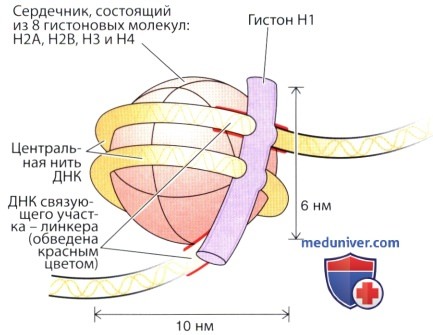
Хроматин состоит преимущественно из скрученных нитей ДНК, связанных с основными белками (гистонами); его структура схематически представлена на рисунке. Главной структурной единицей хроматина является нуклеосома, которая состоит из сердечника, образованного четырьмя типами гистонов (по две молекулы каждого из гистонов Н2А, Н2В, Н3 и Н4), вокруг которого намотаны 166 пар оснований ДНК.
Дополнительный сегмент, состоящий из 48 пар оснований, образует связующее звено (линкер) между соседними нуклеосомами, причем с этой ДНК связаны другие типы гистонов (H1 или Н5). Такая организация хроматина описана под названием «бусы на нити». Негистоновые белки также ассоциированы с хроматином, однако их расположение значительно менее понятно.
Следующим, более высоким, уровнем организации хроматина является волокно толщиной 30 нм. В этой структуре нуклеосомы скручены вокруг оси, причем на каждый виток приходится по шесть нуклеосом, в результате чего образуется 30-нанометровое хроматиновое волокно. Существуют и более высокие уровни скручивания, особенно при конденсации хроматина во время митоза и мейоза.
Характер распределения хроматина в ядре рассматривается как показатель клеточной активности. Как правило, клетки со светлыми ядрами более активны, чем клетки с конденсированными, темными ядрами. В светлоокрашенных ядрах (с небольшим количеством глыбок гетерохроматина) имеется большая поверхность Д Н К, доступная для транскрипции генетической информации. В темноокрашенных ядрах (богатых гетерохроматином) вследствие скручивания ДНК поверхность, доступная для транскрипции, уменьшена.
Эта структура состоит из сердечника, образованного четырьмя типами гистонов (по две молекулы каждого) — Н2А, Н2В, Н3 и Н4 — и одной молекулы гистонов H1 или Н5, расположенных вне нити ДНК.
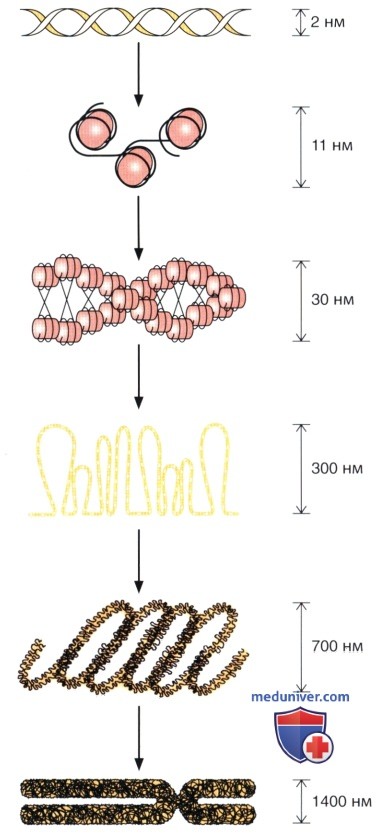
Начиная сверху, изображена двойная спираль ДНК толщиной 2 нм; далее показана ассоциация ДНК с гистонами с образованием нуклеосомных нитей толщиной 11 и 30 нм.
При последующей конденсации формируются филаменты диаметром 300 и 700 нм. Наконец, нижний рисунок показывает метафазную хромосому, в которой выявляется максимально выраженная упаковка ДНК.
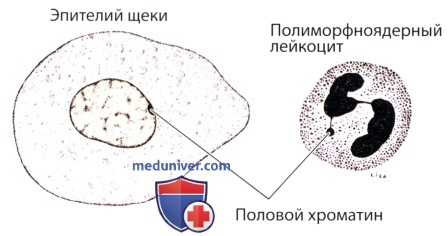
В эпителии половой хроматин имеет вид мелкой плотной гранулы, прилежащей к ядерной оболочке.
В лейкоците для него типична форма барабанной палочки.
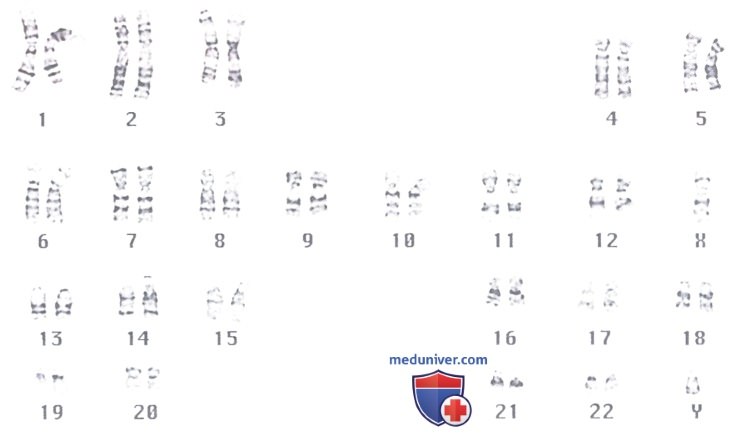
Препарат получен с использованием метода, выявляющего полосы на хромосомах.
Для каждой хромосомы характерен индивидуальный рисунок полос, который облегчает ее идентификацию, а также выявляет связь характера полос с генетическими аномалиями.
Хромосомы группируют в нумерованные пары в соответствии с их морфологическими характеристиками.
Тщательное исследование хроматина ядер клеток млекопитающих обнаружило скопление гетерохроматина, которое часто выявляется в клетках самок, но отсутствует в клетках самцов. Эта глыбка хроматина представляет собой половой хроматин и является одной из двух Х-хромосом, присутствующих в клетках у особей женского пола. Х-хромосома, которая образует половой хроматин, остается плотно скрученной и заметной, а другая Х-хромосома раскручена и невидима. Имеются данные о том, что половой хроматин генетически неактивен.
У особей мужского пола имеется одна Х-хромосома и одна Y-хромосома, которые служат детерминантами пола; Х-хромосома находится в раскрученном состоянии, поэтому половой хроматин не выявляется. В эпителиальных клетках человека половой хроматин имеет вид мелкой гранулы, прикрепленной к ядерной оболочке. Для исследования полового хроматина часто используют клетки, выстилающие внутреннюю поверхность щеки. С этой целью зачастую изучают и мазки крови; в этом случае половой хроматин имеет вид добавочного сегмента в форме барабанной палочки в ядре нейтрофильных лейкоцитов.
Существенный прогресс в изучении хромосом был достигнут после разработки методов, которые включают индукцию деления клеток, остановку митотического деления клеток в метафазе и разрушение (разрыв) клеток. Митоз можно индуцировать фитогемагглютинином (в клеточных культурах) и остановить в метафазе колхицином. Далее клетки погружают в гипотонический раствор, который вызывает их набухание, после чего их распластывают и разрушают между предметным и покровным стеклами. Набор хромосом, полученный из клетки человека, после их окрашивания, представлен на рисунке.
Выделяют половые Х- и Y-хромосомы, остальные же хромосомы обычно группируют на основании их размеров и морфологических характеристик в 22 последовательно пронумерованные пары.
Исследование полового хроматина выявляет генетический пол тех пациентов, у которых по виду наружных половых органов невозможно определить половую принадлежность, как, например, при гермафродитизме и псевдогермафродитизме. Изучение полового хроматина помогает в исследовании других аномалий, затрагивающих половые хромосомы, например синдрома Клейнфельтера, при котором аномалии яичка, азооспермия (отсутствие сперматозоидов) и другие симптомы связаны с наличием хромосомного набора XXY.
- Читать далее "Строение, функции ядрышка"
Оглавление темы "Строение клеток":- Строение, функции хроматина
- Строение, функции ядрышка
- Фазы клеточного деления - митоза
- Фазы клеточного цикла: характеристика, продолжительность
- Апоптоз клеток. Механизмы
- Виды тканей человека. Характеристика
- Формы эпителиальных клеток. Характеристика
- Базальная пластинка и базальная мембрана тканей: гистология, строение
- Межклеточная адгезия и межклеточные соединения в тканях. Виды
- Строение, функции микроворсинок эпителиальных клеток