
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Фазы клеточного цикла: характеристика, продолжительность
Митоз является видимым проявлением клеточного деления, однако в размножении клеток важнейшую роль играют и другие процессы, не столь легко наблюдаемые под световым микроскопом. Главным из них является репликация ДНК. Этот процесс можно проанализировать путем введения в клетку радиоактивных предшественников ДНК (например, [Н] тимидина) и их последующего выявления биохимическими и авторадиографическими методами.
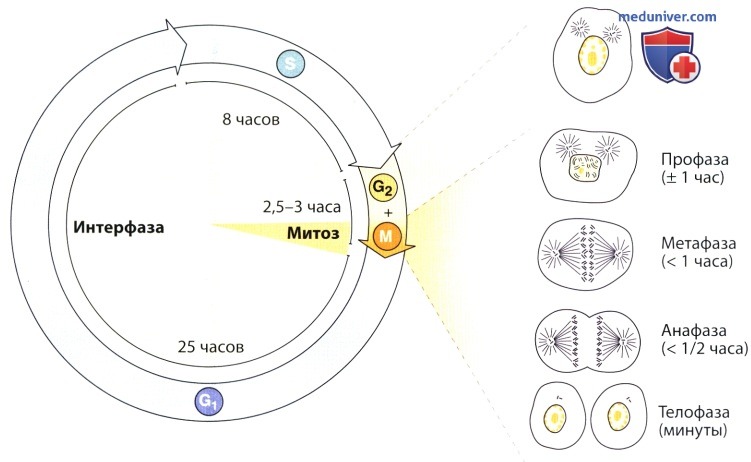
Период, в течение которого происходит репликация ДНК, — интерфаза — не сопровождается заметными при микроскопии явлениями, связанными с клеточным делением. Такое чередование митоза и интерфазы, известное как клеточный цикл, происходит во всех тканях, где имеется обновление клеток. Тщательное исследование клеточного цикла показывает, что его можно разделить на две стадии: митоз, состоящий из четырех фаз (профазы, метафазы, анафазы, телофазы), и интерфазу.
Интерфаза в свою очередь подразделяется на три фазы: G1 (пресинтетическую), S (синтез ДНК) и G2 (после удвоения ДНК). Последовательность этих фаз и приблизительная их продолжительность показаны на рисунке. S-фаза характеризуется синтезом ДН К и началом удвоения центросом с их центриолями.
Во время G1-фазы происходит интенсивный синтез РНК и белков, включая белки, которые регулируют клеточный цикл, при этом объем клетки, ранее уменьшившийся в результате митоза до половины исходного, восстанавливается до своего нормального уровня. В клетках, которые не делятся непрерывно, активность клеточного цикла может прекратиться на время или окончательно. Клетки, находящиеся в таком состоянии (например, мышечные, нервные), описывают как пребывающие в фазе G0.
Регуляция клеточного цикла у млекопитающих осуществляется сложными механизмами. Известно, что культивируемые клетки в отсутствие сыворотки прекращают пролиферацию и останавливаются в G0. В сыворотке крови содержатся высокоспецифические белки — факторы роста — незаменимые компоненты, способствующие делению клеток даже в очень низких концентрациях.
Большинство клеток могут вернуться в цикл из этой фазы, но некоторые из них остаются в G0 в течение длительного времени или даже на протяжении всей своей жизни.
В условиях, неблагоприятных для клетки, цикл останавливается в критической точке, или точке рестрикции (R) в G1.
Когда кчетка преодолевает эту точку рестрикции, она продолжает цикл, проходя через фазу синтеза (S) и G2-фазу, давая в результате митоза (М) начало двум дочерним клеткам, если только не произойдет остановка в другой точке рестрикции (не показана) в G2.

G1-фаза(пресинтетическая) варьирует по длительности в зависимости от многих факторов, включая скорость клеточного деления в данной ткани.
В костной ткани G1-фаза длится 25 ч. S-фаза (синтез ДНК) продолжается около 8 ч.
G2-фаза совместное фазой митоза длится 2,5—3 ч.
Клеточный цикл регулируется также разнообразными сигналами, которые угнетают процесс прохождения клетки по циклу. Повреждение ДНК приводит к остановке клеточного цикла не только в G2, но также и в «контрольной точке» (точке рестрикции) в G1. Остановка в G1 позволяет осуществить устранение поломки (репарацию) ДНК до того, как клетка войдет в S-фазу, в которой происходила бы репликация поврежденной ДНК.
В клетках млекопитающих блокирование цикла в контрольной точке в G1 опосредовано действием белка, называемого р53. Ген, кодирующий р53, в раковых клетках у человека часто изменяется вследствие мутации, тем самым снижается способность клетки осуществлять репарацию поврежденной ДНК. Наследование поврежденной ДНК дочерними клетками приводит к повышенной частоте мутаций и общей нестабильности генома, что может способствовать развитию рака.
Процессы, происходящие в течение G2-фазы, включают накопление энергии, которая будет использоваться во время митоза, синтез тубулина, который подвергнется сборке с образованием микротрубочек митотического веретена, и синтез хромосомных негистоновых белков. В фазе G2 также имеется критическая точка, в которой клетка остается до тех пор, пока не произойдет коррекции всей ДНК, синтезированной с дефектами.
В фазе G2 происходит накопление белкового комплекса — фактора, обеспечивающего созревание, или М-стимулирующего фактора (МСФ, или МPF — maturation promoting factor), который индуцирует начало митоза, конденсацию хромосом, разрыв ядерной оболочки и другие события, связанные с митозом.
Некоторые факторы роста используются в медицине. Примером может служить эритропоэтин, который усиливает пролиферацию, дифференцировку и жизнеспособность предшественников эритроцитов в красном костном мозгу.
Быстрорастущие ткани (например, эпителий кишки) часто содержат митотически делящиеся клетки, чего не наблюдается в медленнорастущих тканях. Увеличенное количество фигур митоза и аномальные митозы в опухолях являются важными характеристиками, которые отличают злокачественные опухоли от доброкачественных. Организм обладает сложными регуляторными системами, которые контролируют репродукцию клеток, либо стимулируя, либо угнетая митоз.
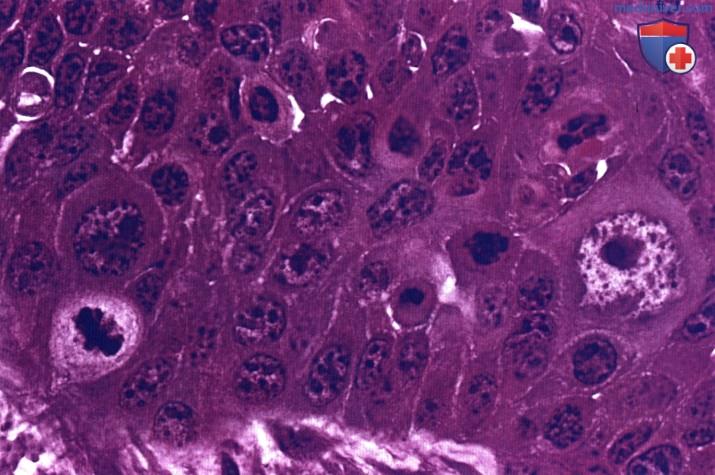
Признаками злокачественности являются увеличение числа митотически делящихся клеток и разнообразие морфологических признаков ядер.
Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
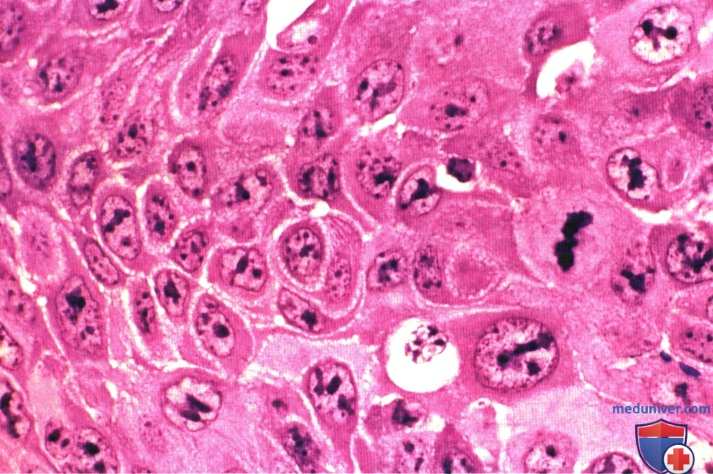
Заметны повышенное количество митотически делящихся клеток и резко выраженное разнообразие морфологических признаков ядер.
Окраска: гематоксилин—эозин. Среднее увеличение.
Нормальную пролиферацию и дифференцировку клеток контролирует группа генов — протоонкогены; нарушения структуры или экспрессии этих генов приводят к развитию опухолей. Измененные протоонкогены содержатся в вирусах, вызывающих опухоли, они, вероятно, имеют клеточное происхождение. Измененная активность онкогенов может возникнуть вследствие нарушения последовательности в молекуле ДНК (мутации), увеличения числа генов (амплификации генов), либо реаранжировки генов, при которой гены перемещаются в участок вблизи активного промотера.
Была установлена связь между измененными онкогенами и развитием некоторых опухолей и гематологических новообразований. Белки, стимулирующие митотическую активность в различных типах клеток, включают фактор роста нервов, эпидермальный фактор роста, фактор роста фибробластов и предшественники фактора роста эритроцитов (эритропоэтина); список этих белков расширяется и быстро растет.
Пролиферация клеток обычно регулируется точными механизмами, которые, когда необходимо, могут стимулировать или задерживать митоз в зависимости от потребности организма. Ряд факторов (например, химические вещества, некоторые виды облучения, вирусные инфекции) способен вызвать повреждение ДНК, мутацию и аномальную пролиферацию клеток, которые обходят нормальные регуляторные механизмы контролируемого роста и приводят к образованию опухолей.
Термин «опухоль», первоначально употреблявшийся для обозначения любой ограниченной припухлости в организме, вызванной воспалением или аномальной пролиферацией клеток, в настоящее время обычно используется как синоним терминов «новообразование, неоплазия» (греч. neos — новый + plasma — образование). Новообразование можно определить как патологическую массу ткани, образованную вследствие нерегулируемой пролиферации клеток.
Новообразования могут быть либо доброкачественными, либо злокачественными в зависимости от имеющихся у них признаков — медленного и неинвазивного роста (доброкачественные) или быстрого роста и выраженной способности врастать в другие ткани и органы (злокачественные). Рак — это распространенный термин, которым обозначают все злокачественные опухоли.
Видео стадии и фазы клеточного цикла клетки
- Читать далее "Апоптоз клеток. Механизмы"
Оглавление темы "Строение клеток":- Строение, функции хроматина
- Строение, функции ядрышка
- Фазы клеточного деления - митоза
- Фазы клеточного цикла: характеристика, продолжительность
- Апоптоз клеток. Механизмы
- Виды тканей человека. Характеристика
- Формы эпителиальных клеток. Характеристика
- Базальная пластинка и базальная мембрана тканей: гистология, строение
- Межклеточная адгезия и межклеточные соединения в тканях. Виды
- Строение, функции микроворсинок эпителиальных клеток