
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Межклеточная адгезия и межклеточные соединения в тканях. Виды
В обеспечении слипания клеток и их сообщения (коммуникации) между собой участвуют несколько структур, связанных с мембраной. Они присутствуют в большинстве тканей, но очень сильно выражены в эпи-телиях, почему они и описаны в настоящей главе. Эпителиальные клетки крайне прочно связаны друг с другом, поэтому для того, чтобы их разъединить, требуются сравнительно существенные механические усилия.
Межклеточная адгезия особенно хорошо выражена в тех эпителиальных тканях, которые подвергаются растяжению и сдавлению (например, в коже). Адгезия частично обусловлена связующим действием семейства трансмембранных гликопротеинов, называемых кадгеринами. Кадгерины утрачивают свои адгезивные свойства в отсутствие Са2+. Интердигитации между складками мембран соседних клеток также способствуют усилению межклеточной адгезии.
Латеральные мембраны многих эпителиальных клеток часто содержат несколько типов модифицированных мембранных участков, известных как межклеточные соединения. Один тип этих соединений обеспечивает механизм коммуникации между соседними клетками. Другие соединения образуют участки адгезии и герметизирующие устройства, предотвращающие ток веществ через пространства между эпителиальными клетками.
В некоторых эпителиях различные межклеточные соединения располагаются в определенном порядке в направлении от апикального полюса клетки к базальному.
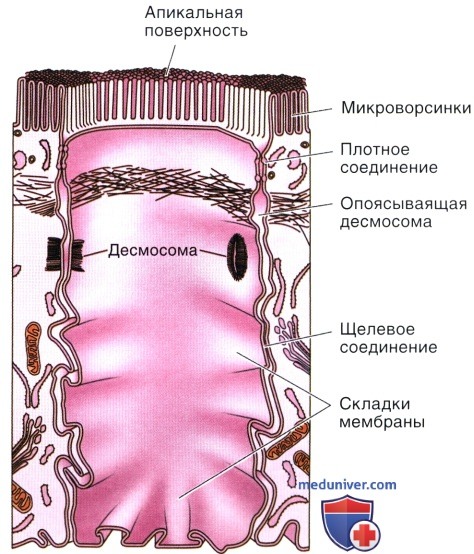
На рисунке показаны три клетки из эпителия кишки. Из средней клетки содержимое удалено для того, чтобы продемонстрировать внутреннюю поверхность мембраны.
В плотном соединении (zonula occludens) наружные пластинки соседних мембран сливаются друг с другом.
Как плотное соединение, так и опоясывающая десмосома (zonula adherens), имеют вид непрерывной ленты, окружающей апикальную часть клетки, тогда как десмосомы и щелевые соединения являются контактами в виде ограниченных участков (пятен).
Складки мембраны и микро ворсинки увеличивают площадь поверхности мембраны и усиливают как обмен веществ через мембрану, так и адгезию между клетками.
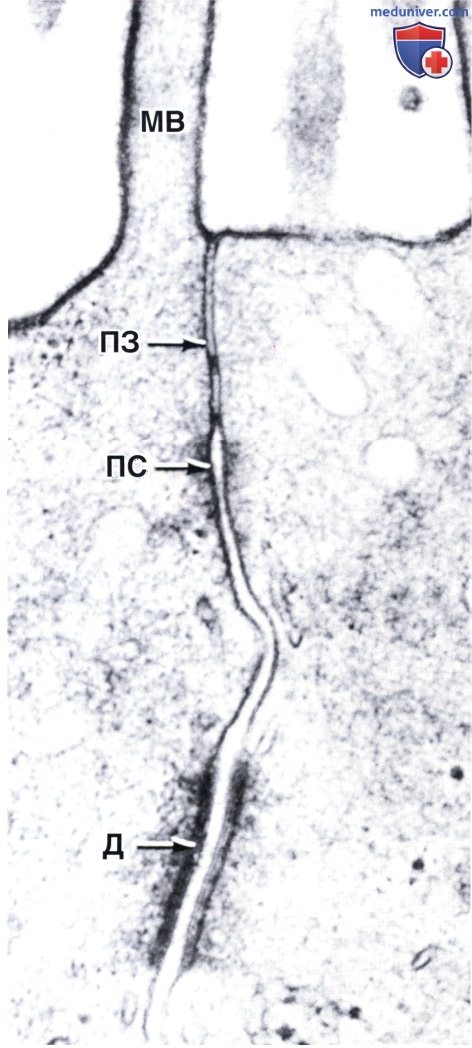
Видны также десмосома (Д) и микроворсинка (МВ). Электронная микрофотография, х80 000.
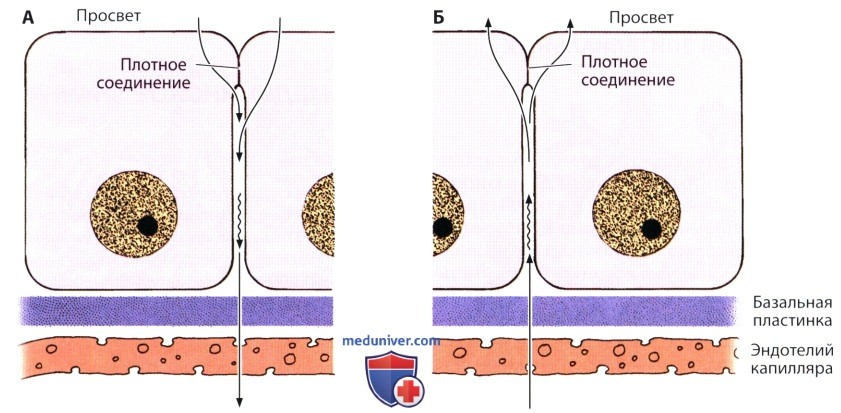
А — направление транспорта — от просвета органа к кровеносному сосуду, как это происходит, например, в желчном пузыре или кишке. Этот процесс известен как всасывание.
Б — транспорт осуществляется в противоположном направлении, как, например, в сосудистом сплетении головного мозга, цилиарном (ресничном) теле или потовой железе. Такой процесс известен как секреция.
Обратите внимание на то, что для поддержания компартментализации (разделения эпителия на части — компартменты), а благодаря ей регуляции распределения ионов, требуется присутствие замыкающих соединений.
Плотные соединения, или пояски замыкания (лат. — zonulae occludens, в ед. числе — zonula occludens), являются наиболее апикально расположенными из всех межклеточных соединений. Латинская терминология дает важную информацию о геометрических характеристиках этого соединения. Слово «поясок» («zonula») отражает тот факт, что это соединение имеет вид полоски, целиком окружающей клетку, а слово «замыкания» («occludens») относится к участкам слияния мембран, которые перекрывают (замыкают) межклеточное пространство.
При исследовании хорошо окрашенных тонких срезов под трансмиссионным электронным микроскопом обнаруживаются участки слияния наружных листков мембран соседних клеток, в результате чего в них образуются пятислойные пласты. В зависимости от типа эпителия можно заметить от одного до нескольких таких участков слияния. При исследовании эпителиальных клеток методом замораживания-скалывания (криофрактографии) можно увидеть анастомозирующие гребешки и канавки, которые образуют сетевидную структуру, соответствующую участкам слияния, выявляемым на обычных тонких срезах.
Между числом гребешков и канавок, или участков слияния, с одной стороны, и проницаемостью эпителия, с другой, существует высокая степень корреляции. Эпителии с одним участком слияния или с их небольшим числом (например, в почечных канальцах проксимального отдела нефрона) более проницаемы для воды и растворов, чем эпителии с многочисленными участками слияния (например, в мочевом пузыре).
Таким образом, главной функцией плотных соединений является формирование «замка», который препятствует перемещению материала в пространстве между эпителиальными клетками (по так называемому парацеллюлярному пути) в обоих направлениях (от апикального края к базальному и от базального к апикальному). Тем самым плотное соединение участвует в образовании двух функциональных отделов (компартментов): апикального, который состоит из полости органа (такой, как, например, просвет кишки) или полости секреторного отдела, и базального, который начинается в области соединений и охватывает подлежащие ткани.
Во многих эпителиях следующим типом соединения, который обнаруживается между клетками, является поясок слипания (лат. — zonula adherens). Это соединение целиком окружает клетку и обеспечивает прилипание (адгезию) одной клетки к соседним. Важным признаком этого соединения является внедрение многочисленных актиновых филаментов в электронно-плотные пластинки материала на цитоплазматических поверхностях мембран в области соединения.
Филаменты принадлежат к сплетению актиновых филаментов, промежуточных филаментов и спектрина вблизи свободной поверхности, известному как терминальная сеть.
Щелевые, или коммуникационные, соединения могут встречаться в любом участке вдоль латеральных мембран эпителиальных клеток. На самом деле щелевые соединения обнаруживаются почти во всех тканях млекопитающих, главным исключением из которых является скелетная мышца.
На обычных трансмиссионных электронных микрофотографиях они выявляются как участки близко (на расстоянии 2 нм) прилежащих друг к другу мембран соседних клеток. На препаратах, полученных методом замораживания-скалывания, эти соединения видны как агрегаты внутримембранных частиц, расположенных в виде округлых скоплений в плазматической мембране.
Структурной единицей щелевого соединения является коннексон. В состав каждого коннексона входят белки щелевых соединений — коннексины, которые образуют группу из шести субъединиц, в центре которой остается гидрофильная пора диаметром 1,5 нм. Коннексоны соседних клеток располагаются на одном уровне, формируя гидрофильный канал между двумя клетками. Каждое щелевое соединение образовано десятками или сотнями стыкующихся между собой пар коннексонов.
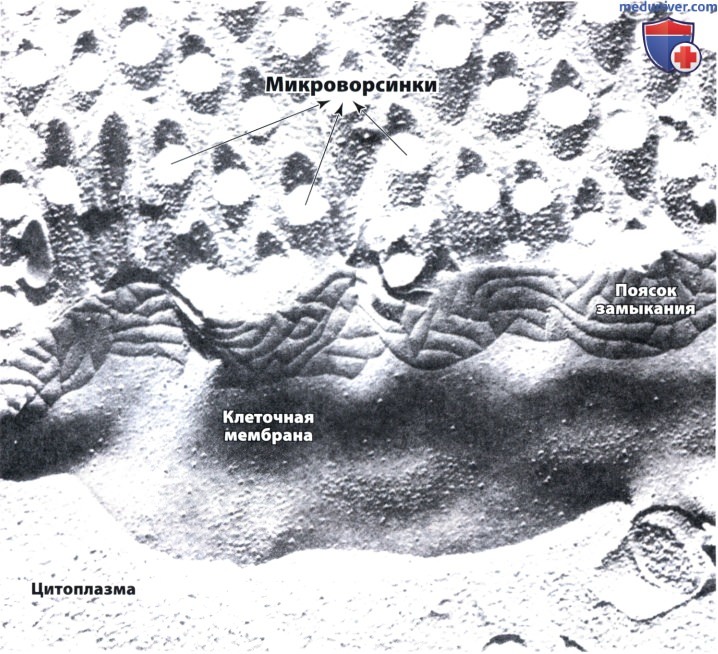
В верхней части видны микроворсинки, сколотые поперечно; в нижней части скол проходит через цитоплазму.
В средней части мембрана расщеплена, и в ней видны бороздки, которые в действительности расположены в липидном слое плазмолеммы.
Эти бороздки участвуют в формировании пояска замыкания. Электронная микрофотография, х 100 000.
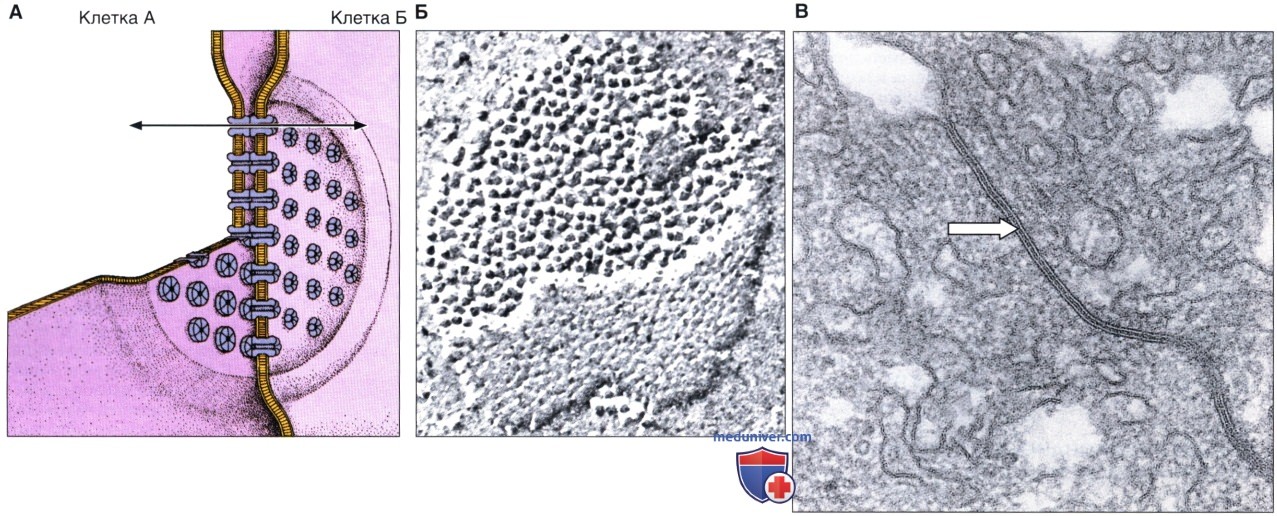
Диаметр канала составляет около 1,5 нм, что ограничивает размер молекул, способных проходить через него. Этим путем из клетки в клетку могут транспортироваться питательные вещества и сигнальные молекулы, при этом не происходит потери материала из-за его попадания в межклеточное пространство.
Б — щелевое соединение — вид на препарате, приготовленном методом замораживания-скалывания. Соединение имеет вид скопления внутримембранных белковых частиц в виде пластинки. х45 000.
В — щелевое соединение (стрелка) клеток печени крысы — вид при использовании трансмиссионной электронной микроскопии. В области соединения мембран прилежащие клетки разделены узким пространством, или щелью, шириной примерно 2 нм, х193 000.
Коннексины принадлежат к семейству родственных белков, которые имеют различное распределение и формируют каналы с различающимися физиологическими свойствами. Щелевые соединения обеспечивают обмен между клетками молекулами с массой < 1500 Дальтон. Сигнальные молекулы, такие, как гормоны, цАМФ и цГМФ (циклический гуанозинмонофосфат), атакже ионы, способны перемещаться через щелевые соединения, благодаря чему клетки во многих тканях функционируют координированно, а не как независимые образования. Типичным примером служат сердечные мышечные клетки (кардиомиоциты), поскольку щелевые соединения между ними в значительной мере ответственны за координированное биение сердца.
Последним типом межклеточных соединений является десмосома (греч. desmos — тяж, soma — тело), или пятно слипания (лат. — macula adherens). Десмосома представляет собой сложную дисковидную структуру на поверхности одной клетки, которой соответствует идентичная структура на поверхности соседней клетки. Клеточные мембраны в этом участке очень прямые и часто располагаются в некотором отдалении друг от друга (на расстоянии >30 нм), что превышает обычный промежуток 20 нм.
С цитозольной стороны мембраны каждой клетки, на небольшом расстоянии от нее, находится округлая пластинка из плотного материала — пластинка прикрепления, в состав которой входят не менее 12 различных белков. В пластинки прикрепления эпителиальных клеток проникают группы промежуточных цитокератиновых филаментов, либо же они заворачивают обратно (наподобие шпильки для волос) и возвращаются в цитоплазму. Поскольку промежуточные филаменты цитоскелета эпителиальных клеток очень прочные, десмосомы обеспечивают устойчивую адгезию между клетками. В неэпителиальных клетках промежуточные филаменты, прикрепленные к десмосомам, состоят не из цитокератина, а из других белков, таких, как десмин или виментин.
В адгезии, которая обеспечивается десмосомами, участвуют белки семейства кадгеринов. В условиях in vitro адгезивные свойства утрачиваются после удаления Са2+.
В зоне контакта между некоторыми эпителиальными клетками и базальной пластинкой часто обнаруживаются полудесмосомы. Эти структуры имеют вид половины десмосомы и прикрепляют эпителиальную клетку к подлежащей базальной пластинке. Однако в десмосомах пластинки прикрепления содержат преимущественно кадгерины, тогда как в полудесмосомах пластинки прикрепления состоят из интегринов, семейства трансмембранных белков, которые являются участками связывания неклеточных макромолекул — ламинина и коллагена IV типа.
- Читать далее "Строение, функции микроворсинок эпителиальных клеток"
Оглавление темы "Строение клеток":- Строение, функции хроматина
- Строение, функции ядрышка
- Фазы клеточного деления - митоза
- Фазы клеточного цикла: характеристика, продолжительность
- Апоптоз клеток. Механизмы
- Виды тканей человека. Характеристика
- Формы эпителиальных клеток. Характеристика
- Базальная пластинка и базальная мембрана тканей: гистология, строение
- Межклеточная адгезия и межклеточные соединения в тканях. Виды
- Строение, функции микроворсинок эпителиальных клеток