
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Схема синтеза порфиринов и их норма
Порфирии относятся к одним из самых интригующих заболеваний человечества. Совершенно разнообразные, часто с нетипичными клиническими проявлениями, эти нарушения обмена порфирина или его предшественника являются результатом аберраций в контролировании процесса биосинтеза гема. Гем необходим для связывания кислорода и его транспортировки (как в форме гемоглобина, так и миоглобина), транспорта электронов (в форме цитохромов) и для активности многофункциональных оксидаз, таких как цитохром Р450. Хлорофилл, магний-хелатный комплекс порфирина, представляет собой другой важный тетрапиррол, имеющий решающее значение для фотосинтеза — специализированной энергозапасающей системы растений, при которой конверсия энергии солнечного света в стабилизированную энергию химических молекул достигается путем последовательности окислительно-восстановительных реакций. Корриновое кольцо, кобальт-хелатный комплекс тетрапиррола, является основной составляющей витамина В12, дефицит которого приводит к развитию пернициозной анемии.
Таким образом, порфирины — это повсеместно необходимые биохимические компоненты живых существ. Биологическое значение порфиринов и их комплексов с железом для процессов метаболизма лежит в их способности действовать в качестве медиаторов окислительных реакций, либо как окислительных компонентов в метаболизме гормонов, лекарственных препаратов и химических веществ окружающей среды, либо в процессах обмена газов, таких как кислород и углекислый газ, между внешней средой и тканями организма.
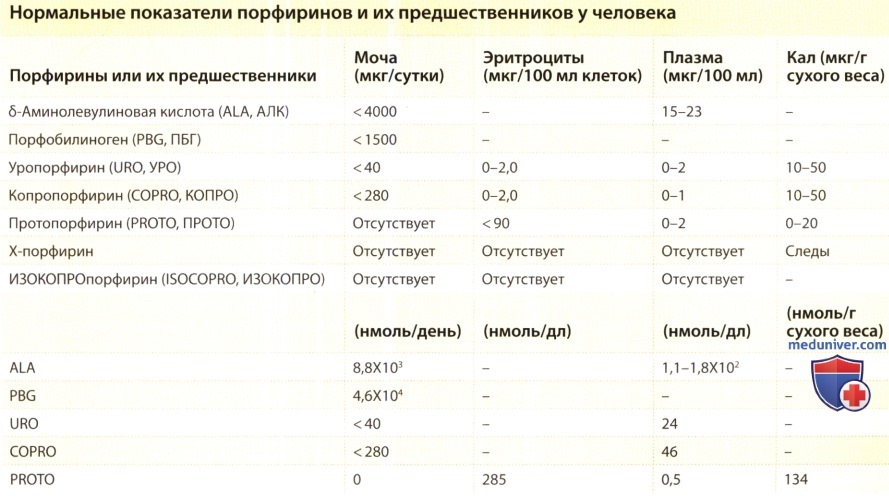
Ежедневный синтез порфиринов и гема в организме человека производится в количествах, достаточных для обеспечения метаболических потребностей. Контроль синтеза гема настолько точен, что, при нормальных условиях, лишь микрограммы (или даже меньше) промежуточных продуктов этого процесса присутствуют в плазме, эритроцитах, моче и кале.
Порфирии — это клинически и генетически гетерогенная группа метаболических заболеваний, являющихся следствием либо наследственной, либо приобретенной дисфункции ферментов, необходимых для биосинтеза гема. Синтез гема контролируется восемью ферментами на протяжении всего процесса биосинтеза. Недостаточная активность семи из этих восьми ферментов может вызвать специфический тип порфирии. Усиление функционирования первого энзима в цепочке (синтетазы δ-аминолевулиновой кислоты) отвечает за развитие Х-сцепленной доминантной протопорфирин (ХСДПП), последней из открытых типов порфирии. Каждый из этих ферментов обсуждается отдельно далее в контексте относящегося к нему типа порфирии.
Биохимически различные порфирии характеризуются определенными моделями аккумуляции и экскреции порфиринов и/или их предшественников. В общем, порфирины, выделяемые при специфическом типе порфирии, являются необратимо окисленными субстратами дефицитного фермента. Эти промежуточные вещества, если они находятся в избыточном количестве, приводят в действие токсические эффекты, которые ответственны за кожные и неврологические симптомы порфирии. Порфирии имеют особенный дерматологический интерес, так как при нескольких их типах обнаруживаются кожные признаки, позволяющие поставить диагноз, исходя лишь из клинической картины.
Основными органами, где происходит синтез гема, являются костный мозг и печень. Гем является составляющей частью ряда белков, среди которых гемоглобин, миоглобин, митохондриальные и микросомальные цитохромы (в том числе цитохром Р450), каталаза, пероксидаза, триптофанпирролаза, простагландин-эндопероксид-синтаза, а также растворимая форма гуанилатциклазы. Почти 85% гема синтезируется в костном мозге, где гем используется для продукции гемоглобина; большая часть остального гема синтезируется в печени для продукции цитохрома Р450, каталазы и разнообразных митохондриальных цитохромов. Гем служит неотъемлемой клеточной составляющей, необходимой для целого ряда метаболических процессов, в первую очередь благодаря его уникальной способности захватывать и освобождать кислород и способствовать транспорту электронов.
Синтез гема регулируется взаимодействием ряда факторов и напрямую зависит от концентрации внутри клеток, а также от потребности клеток в продукции различных гемопротеинов, описанных выше. Многие из них имеют высокую периодичность обновления (от минут до часов), что, в свою очередь, требует постоянно высокого уровня синтеза гема печенью. Например, цитохром Р450, важный мембрансвязанный энзим печени, участвующий в дезинтоксикации и метаболизме лекарственных препаратов, имеет период полураспада в 90-180 минут.
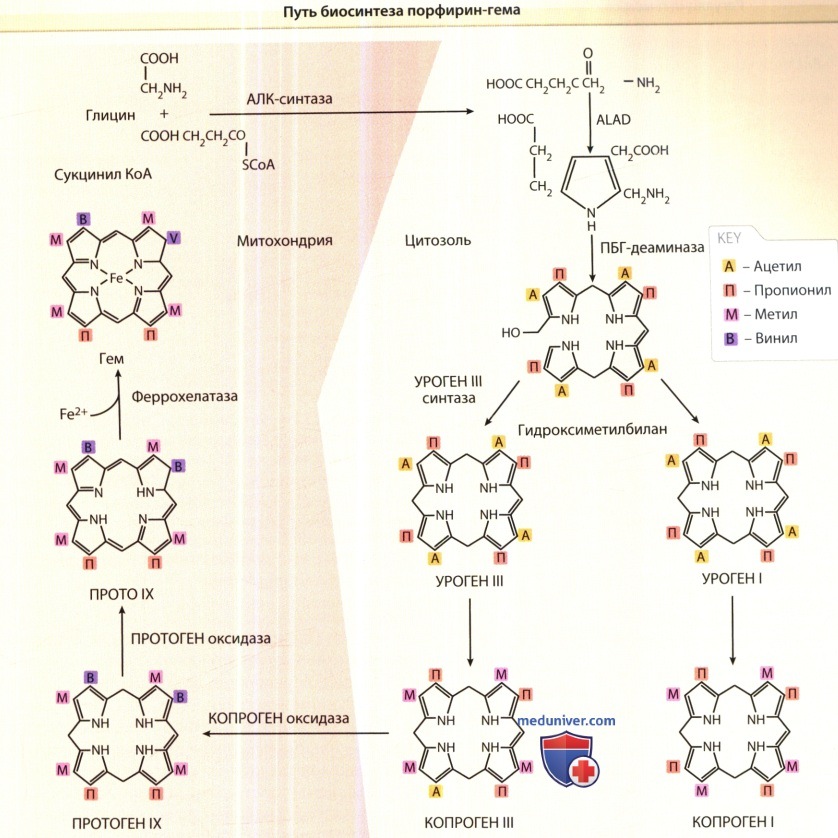
Регуляция синтеза порфирина и гема. Синтез гема начинается в митохондриях клетки, где сукцинат и глицин (отдельные моллекулы глицина и сукцинил КоА) конъюгируются синтетазой δ-аминолевулиновой кислоты и образуют пятиуглеродный аминокетон — гамма-аминолевулиновую кислоту. В качестве кофактора эта реакция требует участия пиридоксаль-5’-фосфата. Регуляция биосинтеза гема (а также способности синтезировать гем вообще) зависит от ряда взаимодействий восьми внутриклеточных ферментов. Первый, а также три последних вовлеченных в процесс фермента, копропорфириноген оксидаза (КПО), протопорфериногеноксидаза (ППО) и феррохелатаза (ФеХ), являются митохондриальными, а остальные ферменты, включая дегидратазу δ-аминолевулиновой кислоты (ДАК), порфобилиногендезаминазу (ПБД), уропорфириногенсинтазу (УПГС) и уропофириногендекарбоксилазу (УПГД), локализованы в цитозоле. Катализатором при распаде гема являются микросомальные ферменты НАДФН-цитохром-С-редуктаза и гемоксигеназа 1 и 2. Гемоксигеназа 1 типа—индикатор оксидативного поражения ткани и сильный антиоксидант. Гем оксигеназа 2 типа—это сенсор килорода. Продукты метаболизма этих ферментов (линейный тетрапирролбиливеодин-IXa и монооксид углерода) также обладают антиоксидантными свойствами.
кДНК каждого из восьми ферментов, которые участвуют в биосинтезе гема наравне с ферментами, отвечающими за катаболизм гема, были клонированы в клетках млекопитающих. Нарушения в регулировании синтеза гема могут быть результатом дефектов ферментов, а это, в свою очередь, может случаться в результате наследственных факторов и/или факторов окружающей среды, что приводит к накоплению в организме одного или более промежуточных продуктов синтеза гема, таких как порфирины или их предшественники. Хотя диагноз порфирии часто может быть выставлен лишь на основании тщательного сбора анамнеза (в том числе, семейного) и осмотра, точная диагностика требует определения: (1) порфиринов и/или их предшественников в моче, кале, плазме или эритроцитах, или (2) остаточной активности специфичных ферментов в процессе биосинтеза гема, или (3) определения мутаций в генах, кодирующих эти ферменты. Современные технологии молекулярной биологии помогли прояснить основы механизмов развития порфирий. Стадии биосинтеза порфиринов и гема и их регуляция обобщены на рисунках ниже.
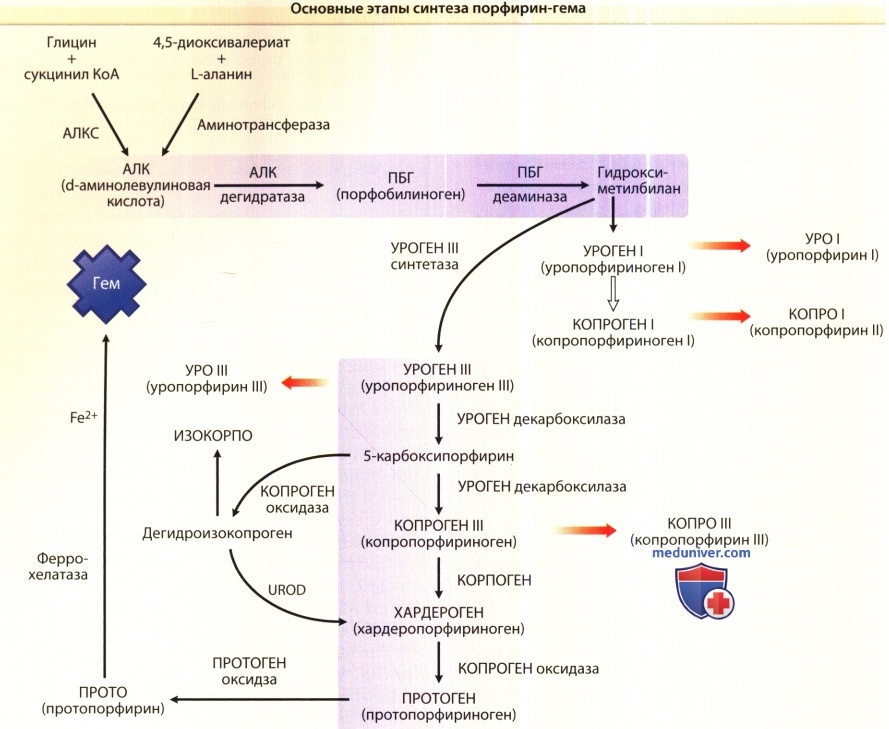
Аминолевуленовая кислота может образовываться из глицина и сукцинилКоА(СКоА), что является основным источником у млекопитающих и катализируется митохондриальным энзимом АЛА синтазой (АЛАС).
Две молекулы АЛА формируют монопирроль порфобилиногена (ПБГ) в реакции, катализированной ферментом АЛА дегидратазой (АЛАД).
Четыре молекулы ПБГ упакованы с помощью ПБГ деаминазы (ПБГД), также известной как гидроксиметилбилансинтаза (ГМБС), в линейный тетрапиррол, гидрокисметилбилан (ГМБ), который может спонтанно зациклироваться и образовывать уропорфириноген (УРОГЕН) 1.
Четыре ацетильные группы УРОГЕНа 1 последовательно декарбоксилируются уропорфириногендекарбоксилазой (УРОД), чтобы образовать копропорфириноген (КОПРОГЕН) 1.
ГМБ также может превращаться в УРОГЕН III с помощью фермента уропрфириноген III синтазы (УРОС), в этой реакии одно из колец монопирроля «опрокидывается», что меняет последовательность концевых цепей.
Ацетильная группы УРОГЕН III последовательно декарбоксилируются с помощью УРОД до КОПРОГЕН III.
КОПРОГЕН III превращается в протопорфириноген (ПРОТОГЕН) IX с помощью фермента копропорфириноген оксидазы (КПОКС), которая оксидативно декарбоксилирует каждую пропионильную группу.
ПРОТОГЕН IX превращается в протопорфирин (ПРОТО) IX с помощью протопорфириногеноксидазы (ППОКС). ПРОТО IX превращается в гем с помощью феррохелатазы (ФЕХ), которая катализирует внедрение в молекулу закис-ных соединений железа.

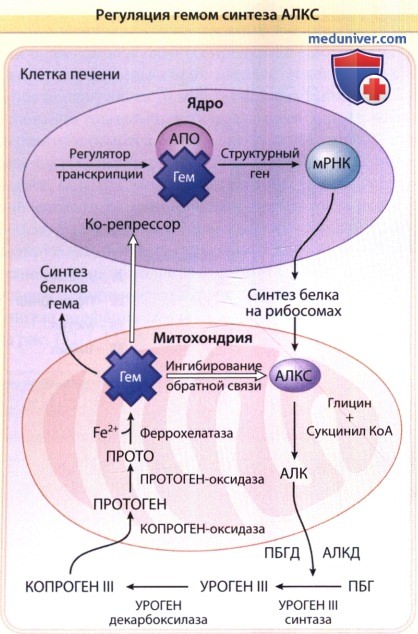
Подавление может являться результатом связывания гема как, например, ко-репрессора, к белку апорепресору (АПО), который, при связывании с гемом, становится функциональным субстратом, блокирующим транскрипцию матричной РНК (мРНК) АЛКС.
Гем также может блокровать транспорт голофермента в митоходрии.
АЛК — δ-аминолевулиновая кислота; АЛКД — дегиратаза δ-аминолевулиновой кислоты; КоА—коэнзим А;
копроген — копропорфириноген; ПБГД—порфобилино-гендеаминаза;
ПРОТО — протопорфирин; протоген — протопорфириноген; уроген—уропорфириноген.
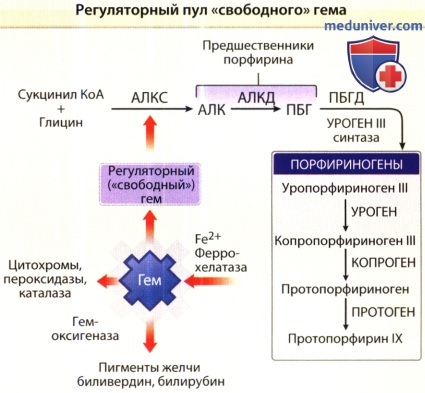
АЛК — δ-аминолевулиновая кислота; АЛКД — дегидратаза δ-аминолевулиновой кислоты;
КоА — коэнзим А; ПБГ—порфобилиноген; ПБГД — порфобилиногендеаминаза;
копроген — копропорфириноген; протоген — протопорфириноген; уроген—уропорфириноген.
- Рекомендуем далее ознакомиться со статьей "Классификация порфирий"
Редактор: Искандер Милевски. Дата публикации: 29.11.2018