
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Внешний путь развития апоптоза
Внешний путь развития апоптоза инициируется рецепторами смерти плазматической мембраны на разных клетках. Рецепторы смерти являются белками семейства рецепторов TNF, содержащими цитоплазматический домен, вовлеченный во взаимодействие белок-белок и называемый доменом смерти, поскольку он необходим для передачи сигналов к запуску апоптоза. (Некоторые белки семейства рецепторов TNF не содержат домена смерти, их функция состоит в активации воспалительного каскада, и их роль в запуске апоптоза не совсем понятна.)
Подробно изучены рецепторы смерти TNF-1 и связанный белок Fas (CD95), но были описаны и другие. Механизм развития апоптоза, индуцированный этими рецепторами смерти, хорошо изучен на примере рецептора Fas, экспрессируемого на многих типах клеток.
Его лиганд FasL экспрессируется на Т-клетках (распознающих собственные антигены и удаляющих аутоагрессивные лимфоциты) и на некоторых цитотоксических Т-лимфоцитах (убивающих инфицированные вирусом и опухолевые клетки).
Когда FasL присоединяется к Fas, три молекулы Fas или более собираются вместе и их цитоплазматические домены смерти формируют участок для связи с белком-адаптером, который содержит Fas-ассоциированный домен смерти (FADD).
В свою очередь, FADD, связанный с рецепторами смерти, через домен смерти присоединяется к неактивной форме каспазы-8 — прокаспазе-8 (и прокаспазе-10 у человека). Таким образом, множество молекул прокаспазы-8 собираются вместе и расщепляют друг друга для генерации активной каспазы-8.
Затем фермент запускает активацию каскада каспаз, активируя другие прокаспазы, и активные ферменты опосредуют фазу, запускающую апоптоз. Этот путь развития апоптоза может быть ингибирован белком FLIP, который присоединяется к прокаспазе-8, но не может ее расщепить и превратить в активную форму, т.к. у него отсутствует протеазный домен.
Некоторые вирусы и нормальные клетки продуцируют FLIP и используют его для защиты от Fas-опосредованного апоптоза.
Внутренний и внешний пути развития апоптоза разные, т.к. при инициации вовлекают совершенно разные молекулы, но эти пути могут быть взаимосвязаны. Например, в гепатоцитах и некоторых других видах клеток сигнал Fas активирует белок ВН3-only (Bid), который затем активирует внутренний путь развития апоптоза.
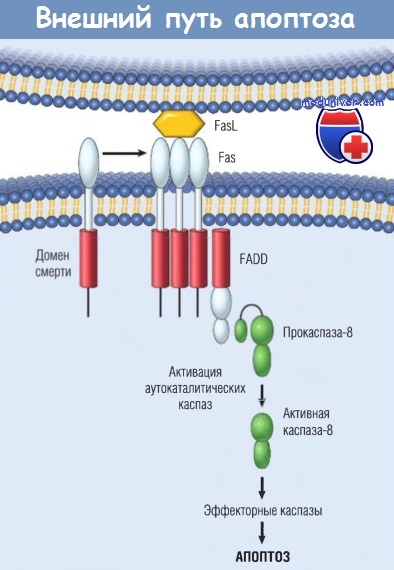
FADD — Fas-ассоциированный домен смерти; FasL — Fas-лиганд.
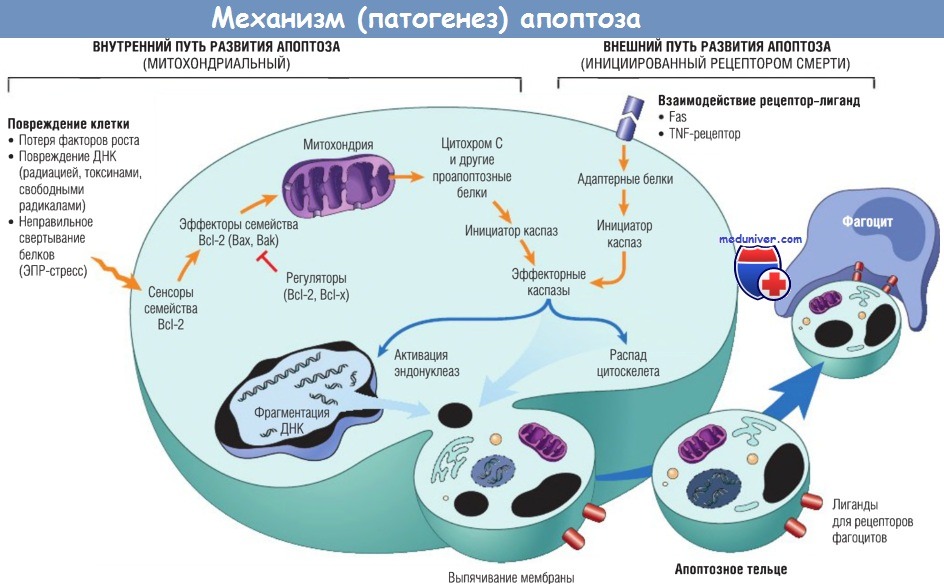
Два пути развития апоптоза отличаются индукцией и регуляцией, но оба заканчиваются активацией эффекторных каспаз.
При внутреннем пути развития апоптоза индукция вовлекает сенсоры и эффекторы семейства Bcl-2, которые индуцируют выход митохондриальных белков.
Показаны также некоторые антиапоптозные белки (регуляторы), которые ингибируют митохондриальный выход и активацию цитохром С-зависимой каспазы во внутреннем пути.
При внешнем пути развития апоптоза инициация рецепторов смерти напрямую вызывает активацию каспаз.
Регуляторы активации каспаз, опосредованные рецептором смерти, не показаны.
TNF — фактор некроза опухоли; ДНК — дезоксирибонуклеиновая кислота; ЭПР — эндоплазматический ретикулум.
- Рекомендуем ознакомиться со следующей статьей "Механизмы запуска апоптоза"
Оглавление темы "Патология клетки":- Ишемическое и гипоксическое повреждение клеток
- Реперфузионное повреждение клетки после ишемии
- Механизмы химического повреждения клетки
- Что такое апоптоз? Причины апоптоза
- Морфологические изменения при апоптозе
- Биохимические признаки апоптоза
- Механизмы апоптоза и их патогенез
- Внутренний путь развития апоптоза
- Внешний путь развития апоптоза
- Механизмы запуска апоптоза