
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Протоонкоген MYC в развитии опухолей
Протоонкоген MYC присутствует фактически во всех клетках эукариотов и принадлежит к группе немедленно реагирующих генов, индуцируемых после получения сигнала к делению покоящейся клеткой. После краткосрочного повышения МУС экспрессия мРНК снижается до исходного уровня.
Молекулярные механизмы функционирования MYC в делящихся клетках остаются до конца не понятыми. Полагают, что участие MYC в канцерогенезе, как и других факторов транскрипции, связано с активацией генов, участвующих в пролиферации. Действительно, некоторые из генов-мишеней MYC — орнитиндекарбоксилаза и циклин D2 — задействованы в клеточной пролиферации.
Однако диапазон активирующего влияния MYC гораздо шире и включает ацетилирование гистонов, снижение адгезивности клеток, увеличение клеточной подвижности, увеличение синтеза белка, снижение протеазной активности и другие изменения метаболизма, обеспечивающие высокий уровень деления клеток. Геномная карта связывающих сайтов MYC содержит тысячи различных участков и эквивалентное количество генов, которые могут регулироваться.
Однако в связи с тем, что генами-мишенями MYC при разных злокачественных опухолях являются разные гены, универсальной концепции участия MYC в канцерогенезе пока нет. Небезынтересно высказанное предположение, что MYC взаимодействует с членами системы ДНК, запускающими процессы репликации, поэтому усиление экспрессии МУС вызывает активацию большего количества генов, участвующих в репликации, чем это необходимо для нормального деления клеток, или дает возможность клеткам уклоняться от точек контроля, что приводит к повреждению генома и накоплению мутаций.
Итак, MYC находится в числе небольшого количества факторов, способных вызывать перепрограммирование соматических клеток в плюрипотентные стволовые клетки; MYC также может усиливать восстановление и дифференцировку клеток.
С одной стороны, активация MYC приводит к пролиферации, а с другой — к апоптозу при отсутствии факторов роста, что было показано на клеточных культурах. Протоонкоген MYC имеет отдельные домены, активирующие как рост, так и апоптоз клеток, однако остается недоказанным, может ли возникать MYC-индуцированный апоптоз in vivo.
В опухолях обнаруживаются персистенция или чрезмерная экспрессия белка MYC в отличие от регулируемой экспрессии при нормальной клеточной пролиферации. Дисрегуляция гена MYC в результате транслокации наблюдается в лимфоме Беркитта, опухоли из В-клеток. MYC амплифицирован в некоторых случаях карцином молочной железы, толстой кишки, легкого и многих других карциномах. Родственные гены N-MYC и L-MYC амплифицированы в нейробластомах и мелкоклеточном раке легкого соответственно.
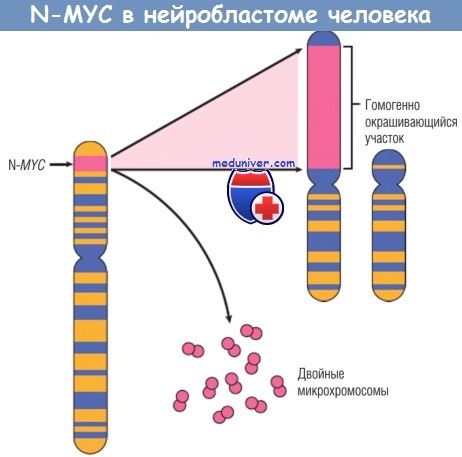
Ген U-MYC в норме локализуется на хромосоме 2р, при амплификации выявляется как добавочные двойные микрохромосомы или гомогенно окрашивающийся участок хромосомы, с которой он интегрировался.
Интеграция обычно происходит с аутосомами 4, 9 или 13.
- Рекомендуем ознакомиться со следующей статьей "Циклины и циклин-зависимые киназы в развитии опухолей"
Оглавление темы "Патофизиология онкологических заболеваний":- Онкоген RAS и его характеристика
- Изменения нерецепторных тирозинкиназ при развитии рака
- Протоонкоген MYC в развитии опухолей
- Циклины и циклин-зависимые киназы в развитии опухолей
- Гены-супрессоры опухоли и их характеристика
- Ген RB и его значение в развитии опухоли
- Ген p53 и его значение в развитии опухоли
- Ген APC и его значение в развитии опухоли
- Ген INK4a/AKF (локус CDKN2A) и его значение в развитии опухоли
- Сигнальный путь TGF-b и его значение в развитии опухоли