
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Гены изменяющие пол. Механизмы детерминации пола у дрозофилы.
Одно из первых прямых доказательств генетической детерминации пола было получено А. Стёртевантом: в 1945 г. им был открыт ген, изменяющий пол и влияющий на развитие первичных половых признаков. В одном из экспериментов по скрещиваниюдрозофил наблюдалось не обычное расщепление по полу (50% самцов и 50% самок), а расщепление, в котором на 62,5% самцов приходилось 37,5% самок, А. Стёртевант показал, что этот феномен обусловлен аутосомным рецессивным геном tra (в схеме скрещивания он обозначен буквой t — трансформатор пола), локализованным в третьей хромосоме дрозофилы. В гомозиготном состоянии этот ген обусловливает развитие фенотипически нормальных, но стерильных самцов из зигот, имеющих две Х-хромосомы. Самец XY, гомозиготный по гену t, является плодовитым.
Если нормальных самок (XX t+t+) скрещивают с самцами, гомозиготными по указанному гену (XY tt), то в первом поколении самки имеют генотип XX t+t, а самцы - XY t+t. В потомстве этих мух происходит следующее расщепление по полу:

Другой пример генетически обусловленной трансформации пола был описан у наездника Habmbracon juglandis. Обычно оплодотворенные яйца развиваются в диплоидных самок, а неоплодотворенные - в самцов. Но иногда самцы появляются и из оплодотворенных яиц. Оказалось, что у Habrobracon juglandis имеется особый ген детерминации пола, и пол зависит от того, находятся ли аллели данного гена в гомо-или гетерозиготном состоянии. Этот ген представлен серией рецессивных аллелей (более 9): Хa, Хb, Хс и т.д. Самки всегда гетерозиготны по этим аллелям, и поскольку они диплоидны, то их генотипы будут: Xa/Xb, Xb/Xc, Xa/Xc и т.д.
Диплоидные самцы, возникающие из оплодотворенных яиц, гомозиготны по одному из серии аллелей: Xa/Xa, Xb/Xb, Xc/XC и тд. Партеногенетические гаплоидные самцы гемизиготны. Если гетерозиготную самку Xa/Xb скрестить с самцом Ха, то в потомстве из оплодотворенных яиц будут развиваться самцы Xa/Xa и самки Xa/Xb. Из-за большого количества аллелей гена, детерминирующего пол у наездника, вероятность случайного образования диплоидного гомозиготного самца весьма мала. Большинство их вследствие причин аналогичных тем, что описаны для самцов-трутней у пчел, погибает еще на стадии личинки, а немногие выжившие оказываются бесплодными.
Семь десятилетий понадобилось генетикам, чтобы ответить на вопрос, какие именно молекулярные механизмы лежат в основе зависимости пола дрозофилы от баланса Х-хромосом и аутосом. В настоящее время гены половой детерминации, идентифицированы; известно также, на каких этапах развития они проявляют свою активность.
Показано, что пол у дрозофилы определяется генами: Sex-lethal (Sxl), transformer (tra), transformer-2 (tra-2), doublesex (dsx) M и F, intersex (ix), sisterless (sis) а и b, daughterless (da).
Установлено, что многоступенчатый процесс половой детерминации начинается с образования комплекса из белковых продуктов генов sis-a и sis-b, расположенных в Х-хромосоме, и аутосомного гена da. Такое различие в локализации названных генов и определяет дальнейшую специфику транскрипции ключевого в определении пола у дрозофилы гена Sxl. Количество продуктов генов sis-a и sis-b находится в прямой зависимости от числа Х-хромосом у эмбриона: одна доза у особей Х/2А и две - у 2Х/2А. В то же время количество продукта гена da и у тех, и у других составляет две дозы вследствие одинакового числа аутосом у самцов и самок.
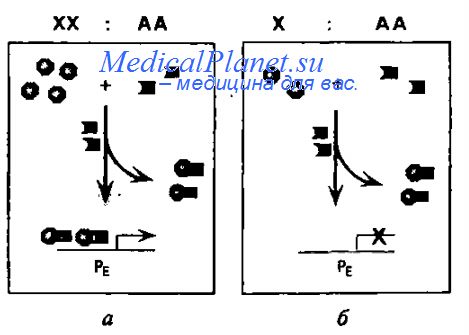
Таким образом, в белковом комплексе sis/da названные компоненты находятся в соотношении 1:2 у эмбрионов Х+Y/2A и 1:1 у эмбрионов 2Х/2А. Именно этот белковый комплекс взаимодействует с регуляторным районом гена Sxl, в состав которого входят ранний (РЕ) и поздний (Pl) промоторы, стимулирующие транскрипцию гена Sxi. Наличие большого числа комплексных молекул sis/da у эмбриона 2Х/2А вызывает активацию раннего промотора (РЕ) на стадии бластодермы, что в конечном итоге приводит к формированию фенотипа самки. У эмбрионов X+Y/2A и Х/2А, имеющих в генотипе единственную Х-хромосому, число комплексных молекул уменьшено вдвое, поэтому активация раннего промотора не происходит и ген Sxl не включается.
На следующих стадиях развития дрозофилы, при активации позднего промотора (PL) продукт гена Sxl, содержащего 8 экзонов, взаимодействует поэтапно с генами tra и dsx. Результаты этого взаимодействия зависят от соотношения Х/А. Так у эмбрионов 2Х/2А ген Sxl (в его составе экзоны 1, 2, 4—8) кодирует функционально полноценный белок. Он в свою очередь вступает во взаимодействие с геном /га, который в комплексе с продуктом гена /га 2 обеспечивает образование специфической для самок РНК гена dsxF. Продукт этого типа вовлекает в цепь формирования пола дрозофилы еще один ген - ix. Именно за счет белковых продуктов генов dsxF и ix на заключительных этапах инактивируются многие гены, которые могли бы репрессировать формирование фенотипа самки.
Напротив, у эмбрионов X + Y/2A в ходе образования первичного транскрипта гена Sxl вслед за экзонами 1 и 2 считывется экзон З, на котором последующая трансляция заканчивается из-за большого числа входящих в его состав стоп-кадров UGA. Усеченный продукт гена Sxl обусловливает специфическое считывание гена tra, в результате чего белок tra тоже оказывается нефункциональным. Это событие определяет сразу два момента в детерминации пола: при отсутствии функционального продукта /га-гена не синтезируется и полноценный продукт гена tra 2, а кроме того, в этих условиях с альтернативного набора экзонов гена dsx считывается белок dsxM, репрессирующий развитие самок. В результате формируются особи с фенотипом самиа.
Целый ряд белков у дрозофилы присущ особям одного пола и не синтезируется у другого. Так, например, для самок характерно наличие специфических белков желтка и яйцевой оболочки (хориона).
Таким образом, в каскад строго иерархических взаимодействий, определяющих детерминацию пола у дрозофилы, вовлечены гены, локализованные как на Х-хромосоме, так и на аутосомах. Последовательная активация либо репрессия этих компонентов каскада в значительной степени обеспечивается альтернативным сплайсингом первичных РНК-транскриптов.
- Читать далее "Пол у человека. Детерминация пола у человека. Роль Y хромосомы в детерминации пола."
Оглавление темы "Пол и гены. Репликация генетического материала.":1. Пол. Дифференцировка пола. Основные виды детерминации пола.
2. Хромосомная детерминация пола. Гинандроморфизм.
3. Баланс полов. Балансовая теория Бриджерса.
4. Гены изменяющие пол. Механизмы детерминации пола у дрозофилы.
5. Пол у человека. Детерминация пола у человека. Роль Y хромосомы в детерминации пола.
6. Аутосомные гены в детерминации пола. Вторичная детерминация пола у человека.
7. Этапы вторичной детерминация пола. Стадии вторичной детерминации пола у человека.
8. Зависимые от пола признаки. Отграниченные от пола признаки.
9. Репликация генетических молекул. Полуконсервативная репликация ДНК и хромосом.
10. Типы репликации геномов. Инициация репликации генома.