
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Зависимые от пола признаки. Отграниченные от пола признаки.
Поскольку генотип всегда выступает как единая, целостная система, то взаимодействие генов, контролирующих механизм дифференцировки пола и иные функции организма, наблюдается нередко. Одним из примеров такого взаимодействия служат признаки, зависимые в своем проявлении от пола и ограниченные полом. Гены, контролирующие такие признаки, могут находиться как в половых хромосомах, так и в любой из аутосом, и проявляются только у одного пола или имеют разную степень проявления у особей противоположного пола.
Например, ген раннего облысения имеет различное проявление у мужчин и женщин. У мужчин этот ген действует как доминантный, у женщин - как рецессивный, поэтому гетерозиготные женщины не проявляют данного признака, а в гомозиготном состоянии он выражен у женщин слабее, чем у мужчин. Высокий уровень мужских половых гормонов определяет доминирование гена рогатости у самцов-баранов, в то время как у самок доминирует ген комолости. Еще одним примером зависимости проявления от пола может служить аутосомный доминантный ген, вызывающий у самцов аквариумной рыбки (сиамского петушка) развитие больших плавников. У самок этот ген не проявляется.
Проявление генов, ограниченных полом, всегда необходимо рассматривать в контексте реализации генотипа как системы в условиях сложного организма высших животных. Помимо генов, ответственных за развитие вторичных половых признаков, которые в норме проявляются только у одного из полов, а у другого могут присутствовать, но не экспрессируются, функциональную активность целого ряда других генов определяет гормональный профиль организма. Так, у быков есть гены, контролирующие продукцию молока и его качественные особенности (жирность, содержание белка и др.), но у быков и их сыновей эти гены «молчат», функционируют они только у их дочерей. Гены, определяющие различия пород по яйценоскости есть как у кур, гак и у петухов, но проявляются они естественно только у самок. Какие из генов есть в наличии у самца можно предполагать только по продуктивности его женских предков и потомков.
В начале данного раздела, уже говорилось о том, что половой диморфизм - основа для воспроизводства потомства. Особь развивается из половой клетки, обладающей потенцией к формированию новых половых клеток и множества соматических клеток. Цикл полового размножения охватывает период от момента появления первичных половых клеток до их образования в следующем поколении. У большинства видов животных и растений в процессе полового размножения происходит слияние мужской и женской гамет, причем строго внутривидовое. Одним из главных механизмов, обеспечивающих этот феномен, является точное соответствие числа и архитектоники хромосом женских и мужских половых клеток, а также сродство цитоплазмы яйцеклетки и ядра сперматозоида.
Партеногенез — тип полового размножения, при котором женские половые клетки развиваются без оплодотворения. Для ряда видов партеногенез — нормальное явление (например, для низших ракообразных, коловраток, перепончатокрылых), но иногда у видов, обычное размножение которых связано с оплодотворением, отдельные яйцеклетки начинают развиваться партеногенетически. Партеногенез обеспечивает воспроизводство при редких контактах разнополых особей и возможность резкого увеличения численности. У некоторых насекомых и ракообразных известен также личиночный партеногенез (педогенез), который служит механизмом, компенсирующим недостаточную плодовитость взрослых особей.
Выделяют следующие естественные формы партеногенеза:
1) амфитокия (дейтеротокия) - из неоплодотворенных яиц развиваются особи обоих полов (известна у некоторых тлей);
2) арренотокия — нео плодотворен ные яйца развиваются только в самцов, а для возникновения самок необходимо оплодотворение (встречается у видов с гаплоидно-диплоидным определением пола - перепончатокрылых насекомых и некоторых членистоногих);
3) телитокия — из неоплодотворенных яиц развиваются только самки (характерна для некоторых тлей, а также ящериц). Во всех случаях телетокия контролируется внутриклеточными эндосимбионтами, которые обусловливают нежизнеспособность самцов.
Как единственная форма размножения партеногенез практически не существует, так как он либо чередуется с половым размножением, либо является принадлежностью отдельных рас. Из позвоночных естественный партеногенез характерен только для пресмыкающихся и индеек. Известно, что до 40% отложенных индейками неоплодотворенных яиц могут начать развиваться, но только в редких, единичных случаях это развитие доходит до конца. Все выведенные таким способом птенцы оказываются самцами. Партеногенез у индеек, скорее всего, представляет собой отклонение от нормального размножения посредством оплодотворения. В отличие от них, у трех рас ящерицы Lacerta saxicola, состоящих из одних самок, партеногенез представляет собой естественный способ размножения.
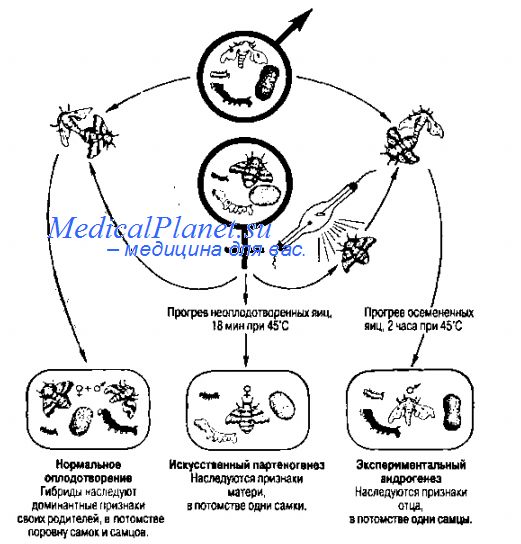
В 1886 г. русский зоолог А.А. Тихомиров показал, что обрабатывая яйца морского ежа хлороформом или стрихнином, можно стимулировать начальные стадии их развития; более того, воздействуя раствором серной кислоты на яйца шелкопряда, ему впервые удалось вызвать искусственный партеногенез. В дальнейшем именно этот объект был использован выдающимся отечественным генетиком БЛ. Астауровым для разработки эффективной технологии партеногенстического размножения шелкопряда в производственных условиях. В соответствии с данной технологией, не прошедшие мейоз и неоплодотворен ные яйца подвергают короткому нагреванию до 46 °С, одновременно активируя и диплоидизируя их (нет редукционного деления и кроссиноговера, и яйца содержат обе половые хромосомы гетерогаметной матери -ZW). Потомство состоит из одних самок. Этот метод амейотического партеногенеза, разработанный в 40-е гг., представляет ценность тем, что потомки в точности воспроизводит генотип матери, и можно быстро и в большом количестве размножать ценный линейный материал.
Такой амейотический партеногенез наблюдается у реснитчатых и круглых червей, коловраток, моллюсков, некоторых видов насекомых, у карпа Carassius auratus gibelio.
Для практического шелководства наиболее выгодным является получение только самцов, так как из их коконов выхолит более длинная шелковая нить. Получение чисто мужского потомства у тутового шелкопряда достигается с помощью экспериментального андрогенеза (термин предложен М. Ферворном в 1892 г.). Андрогенез - развитие организма из яйца с инактивированным ядром, оплодотворенного нормальным спермием, по сути — мужской партеногенез; в природе встречается редко (у некоторых растений и перепончатокрылых насекомых), искусственный андрогенез применяется в селекционно-генетических целях. Таким образом, БЛ. Астауров, вместе со своими сотрудниками разработал способ регуляции пола потомства, одновременно позволяющий с большой точностью исследовать взаимоотношения ядра и цитоплазмы в генетических процессах.
В. А. Струнников разработал метод мейотического партеногенеза, при котором мейоз протекает нормально, а двухчасовым снижением температуры до —{5—11) °С стимулируется развитие неоплодотворенных яиц и диплоидизация во время первого деления. В результате партеногенетическое потомство состоит из одних самцов.
Таким образом, кроме амейотического механизма диплоидизации при партеногенезе, существует мейотический, который, как показано в целом ряде исследований, представлен несколькими вариантами. Диплоидная яйцеклетка может образоваться:
1) при сохранении конъюгации гомологов и кроссинговера, но при отсутствии редукционного деления. Такой способ размножения присущ малому числу партеногенети-чески размножающихся видов, среди которых в основном - насекомые. В эту же группу должны быть включены спонтанные триплоиды у земноводных и человека;
2) при отсутствии эквационного деления. Эта группа более многочисленна и представлена как видами, способными к случайному партеногенезу или дающими редкое триплоидное потомство (включая мышей, крыс и человека), гак и размножающимися партеногенетически или гиногенетически. Гиногенез - форма партеногенеза, при которой спермий не сливается с женским пронуклеусом, а лишь проникает вяйцеклетку и стимулирует ее дробление (известен у некоторых круглых червей, рыб, земноводных и многих покрытосеменных растений), В случае естественного гиногенеза число хромосом у развивающихся особей нормальное, диплоидное. Но искусственный гиногенез часто связан с гаплоидией, что приводит к снижению жизнеспособности зародышей;
3) при слиянии двух гаплоидных ядер: либо гаплоидного пронуклеуса с ядром второго направительного тельца, либо ядра первого направительного тельца с женским пронуклеусом. Существенно, что невозможно различить в эксперименте: отсутствие редукционного деления при наличии кроссинговера и слияние пронуклеуса с ядром первого направительного тельца, а также отсутствие эквационного деления и слияние пронуклеуса с ядром второго направительного тельца;
4) при удвоении хромосом гаплоидного пронуклеуса. Процесс приводит к гомозиготности потомства по всем локусам, что, по-видимому, и является причиной весьма отраниченной распространенности этого механизма у партеногенетических видов;
5) из клеток, в которых произошло премейотическое удвоение, не сопровождающееся цитокинезом. Такое удвоение может привести либо к образованию двуядерного ооцита (например, у лягушек), либо - к формированию тетраплоидного ядра ооцита (например, у планарий),
В настоящее время показано, что искусственный партеногенез возможен практически у всех видов животных, необходим только соответствующий подбор факторов и условий, стимулирующих диплоидность их яйцеклеток. Однако млекопитающие — единственный класс позвоночных, который вследствие геномного импринтинга имеет функциональные различия материнского и отцовского геномов. Результатом этого феномена является ранняя гибель искусственно полученных партеногенетических эмбрионов. Так, в эксперименте было установлено, что диплоидные партеногенетические эмбрионы мышей погибают на преимплантанионных стадиях развития или вскоре после имплантации. С помощью трансформирующего ростового фактора удается пролонгировать развитие партеногенетических эмбрионов и на постимплантационных стадиях.
- Читать далее "Репликация генетических молекул. Полуконсервативная репликация ДНК и хромосом."
Оглавление темы "Пол и гены. Репликация генетического материала.":1. Пол. Дифференцировка пола. Основные виды детерминации пола.
2. Хромосомная детерминация пола. Гинандроморфизм.
3. Баланс полов. Балансовая теория Бриджерса.
4. Гены изменяющие пол. Механизмы детерминации пола у дрозофилы.
5. Пол у человека. Детерминация пола у человека. Роль Y хромосомы в детерминации пола.
6. Аутосомные гены в детерминации пола. Вторичная детерминация пола у человека.
7. Этапы вторичной детерминация пола. Стадии вторичной детерминации пола у человека.
8. Зависимые от пола признаки. Отграниченные от пола признаки.
9. Репликация генетических молекул. Полуконсервативная репликация ДНК и хромосом.
10. Типы репликации геномов. Инициация репликации генома.