
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Типы репликации геномов. Инициация репликации генома.
Репликация начинается с того, что в определенной точке происходит разъединение двойной спирали и образование одноцепочечных участков ДНК, которые служат матрицей для синтеза новой цепи. Участок, в котором в данный момент времени происходит синтез ДНК, называют вилкой репликации. Описано три типа репликации геномов.
1. Репликация бактериальных и вирусных кольцевых геномов начинается с определенной точки и идет в противоположных направлениях, т.е. у бактерий и вирусов существует одна точка начала репликации и две репликационные вилки. Реплицирующаяся хромосома напоминает по структуре греческую букву сигма. По завершении репликации сигма-типа образуются две кольцевые молекулы.
2. У некоторых вирусов (например, у бактериофага X) и при амплификации ДНК генов рРН К в оогенезе у амфибий в одной цепи их кольцевой хромосомы происходит разрыв фосфодиэфирной связи. Затем к свободному 3'-концу разорванной цепи начинают присоединяться нуклеотиды, эта цепь растет, а кольцевая цепь служит матрицей. По мере роста разорванной цепи ее 5'-конец постепенно смещается, и начинается построение цепочки, комплементарной этому участку. Образующаяся структура похожа на греческую букву сигма. Такой тип репликации называют «катящимся кольцом» или типом. Вновь синтезированный «хвост» в определенных точках разрезается, и по завершении оштого цикла репликации образуется одна кольцевая молекула и одна линейная. Длина образующегося «хвоста» иногда может в несколько раз превышать длину окружности кольцевой молекулы.
3. Линейные хромосомы (у некоторых вирусов и эукариот) начинают реплицироваться в одной или нескольких точках, две вилки репликации движутся в противоположных направлениях. По завершении репликации образуются две линейные молекулы.
Участок генома в пределах которого репликация начинается и заканчивается, называется репликоном. Геномы прокариот удваиваются целиком, водном цикле репликации, следовательно, их геномы представляют собой один репликон. В геномах эукариот точек начала репликации множество (несколько сотен или тысяч). Репликация ДНК начинается одновременно во многих точках, следовательно, геном представлен множеством репликонов.
Как в любом матричном процессе, в репликации можно выделить три этапа: инициацию, элонгацию и терминацию.
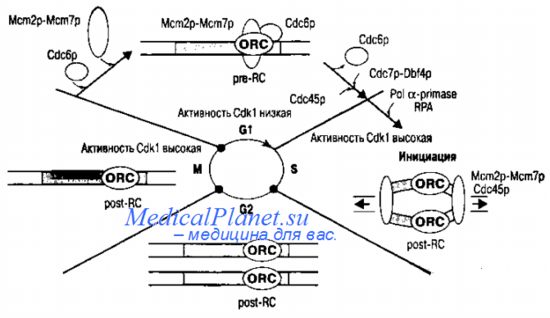
Инициация репликации генома.
Инициация репликации включает формирование репликационной вилки и синтез РНК-праймера. В этом процессе участвует большое число белков и ферментов. Инициирующие белки должны выполнить, по крайней мере, три основные функции: 1) облегчить раскручивание молекул ДНК и ее локальную денатурацию в области начала репликации; 2) обеспечить связь белков и ферментов, участвующих в репликации, с точками начала репликации; 3) обеспечить координацию клеточного цикла и процессов репликации. Для инициации репликации у эука-риот, в отличие от прокариот, связывания инициирующих белков с точками начала репликации недостаточно. Достижение компетентности в данном случае -сложный многоэтапный процесс.
Инициация репликации происходит в строго определенных участках. Выделены и определены последовательности нуклеотидов в точках начала репликации у кишечной палочки Е. coii, многих фагов и плазмид, у дрожжей, млекопитающих и некоторых вирусов эукариот.
У Е. coli этот сайт представляет собой участок ДНК размером 245 нуклеотидов, состоящий из серии 9- и 13- нуклеотидных повторов. Область oriC у бактерий очень консервативна, хотя есть виды, у которых она не обнаружена. Процесс инициации начинается с присоединения к хромосоме белка DnaA. Это приводит к разделению цепей и способствует работе основного расплетающего белка - геликазы (DnaB). В решении топологических проблем, связанных с разделением цепей двойной спирали, участвует и фермент гираза. С образовавшейся одноцепочечной ДНК связываются белки SSB (от англ. single strand binding), которые стабилизируют вилку репликации. Фермент праймаза синтезирует РНК-праймеры на лидирующей и отстающей цепях,
Размер и структура элементов, обеспечивающих начало репликации у эукариот и прокариот, различны. Общим для всех сайтов начала репликации является их обогащенность АТ-парами. По-видимому, это необходимо для обеспечения локальной денатурации, поскольку АТ-пары образуют только две водородные связи.
События, происходящие при инициации репликации у эукариот и связи ее с клеточным циклом, лучше всего изучены у дрожжей. Рассмотрим инициацию репликации и клеточный цикл у дрожжей Saccharomyces cerevisiae. На стадии G1, когда активность циклин-зависимой киназы Cdkl низка, формируется пре-репликационный комплекс, в состав которого входят шесть белков комплекса ORC (ORC1-6) и белки Cdc6 и Mem. Высоко консервативные белки, составляющие комплекс ORC специфически связываются с точками начала репликации и служат основой для присоединения других инициирующих белков Cdc6 и Mem. При переходе от стадии G1 к стадии S активность Cdkl возрастает и Cdc6p покидает комплекс. На его место «встает» белок Cdc45. В этой перестройке комплекса, необходимой для активации точки начала репликации в течение стадии S, принимает участие белок Cdc7-Dbf4-киназа.
После инициации репликации пререпликационный комплекс превращается в пост-репликационный, он состоит только из белков ORC, связанных с хроматином. Этот комплекс сохраняется до конца митоза, когда активность Cdk l падает. Образование нового пре-репликационного комплекса становится возможным только в следующей стадии GI. Таким образом, в течение одного клеточного цикла происходит лишь один цикл репликации. Белки ORC остаются связанными с точкой начала репликации, другие компоненты пре-репликационного комплекса или покидают его, или становятся частью вилки репликации. Например, белки Mcm2p-Mcm7p, по-видимому, функционируют как репликативная геликаза. У всех изученных эука-риот схема событий и белки, участвующие в инициации, сходны. Однако есть и некоторые отличия. Так, у некоторых организмов (другой вид дрожжей, дрозофила, ксенопус)для присоединения Мсm2р-Мсm7р к хроматину необходим дополнительный белок Cdt 1.
У дрожжей белки ORC остаются связанными с хроматином на всех стадиях клеточного цикла, а у позвоночных во время митоза они отделяются от хроматина и вновь соединяются с ним только в стадии G1. До сих пор не ясно, как репликационная машина (ДНК-полимераза-праймаза и репликационный белок А) связывается с точкой начала репликации, как части инициирующего комплекса (Mcm2p-Mcm7p и Cdc45p) преобразуются в компоненты вилки репликации. Гены, кодирующие основные белки, участвующие в инициации репликации ДНК у человека, приведены в таблице.
Разделение двойной спирали происходите помощью ДНК-геликазы и реплика-ционного белка RPA (от англ. - replication protein А). Репликационный белок А, состоящий из трех полипептидов, связывается с одноцепочечный ДНК, таким образом он выполняет ту же функцию, что и SSB-белки у кишечной палочки. Затем а-ДНК-полимераза-праймаза синтезирует короткие (длиной примерно 30 п.н.) РНК-праймеры на лидирующей и отстающей цепях. После этого происходит замена альфа-полимеразного комплекса на комплекс 5-ДНК- полимеразы - основного фермента репликации ДНК у эукариот.
- Вернуться в оглавление раздела "Генетика."
Оглавление темы "Пол и гены. Репликация генетического материала.":1. Пол. Дифференцировка пола. Основные виды детерминации пола.
2. Хромосомная детерминация пола. Гинандроморфизм.
3. Баланс полов. Балансовая теория Бриджерса.
4. Гены изменяющие пол. Механизмы детерминации пола у дрозофилы.
5. Пол у человека. Детерминация пола у человека. Роль Y хромосомы в детерминации пола.
6. Аутосомные гены в детерминации пола. Вторичная детерминация пола у человека.
7. Этапы вторичной детерминация пола. Стадии вторичной детерминации пола у человека.
8. Зависимые от пола признаки. Отграниченные от пола признаки.
9. Репликация генетических молекул. Полуконсервативная репликация ДНК и хромосом.
10. Типы репликации геномов. Инициация репликации генома.