
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Этапы вторичной детерминация пола. Стадии вторичной детерминации пола у человека.
Под контролем другого гормона (андрогена) - тестостерона в ходе вторичной детерминации из вольфовых протоков формируются мужские внутренние половые протоки: средняя часть протоков удлиняется и преобразуется в семявыносящие канальцы. Кроме того, под влиянием тестостерона индуцируется развитие семенных пузырьков и придатка яичка (эпидидимиса). В мочеполовом синусе тестостерон превращается в 5а-дегидротестостерон, при участии которого формируются наружные половые органы; половой член, простата и мошонка. Оба гормона оказывают как местное, гак и общее воздействие, маскулинизируя экстрагенитальные ткани-мишени и обусловливая половой диморфизм центральной нервной системы, внутренних органов и размеров тела.
В случае нарушения биосинтеза андрогенов развиваются отклонения от нормального мужского фенотипа, степень которых может варьировать от легкой гипоспадии (низко расположенного наружного отверстия мочеиспускательного канала) и/или крипторхизма до выраженного женского фенотипа. Сущестпенно, что для нормального развития мужских половых органов необходим не только достаточный уровень андрогенов, но и нормально функционирующие андрогено-вые рецепторы. В отсутствие рецепторов развиваются различные варианты так называемого синдрома нечувствительности к андрогенам (AIS). Андрогеновый рецептор кодируется геном AR, локализованным в Х-хромосоме влокусе ql 1. В его восьми экзонах обнаружено более 200 различных мутаций, среди которых наиболее часто встречаются точковые.
Замена даже одного нуклеотида может приводить к серьезному нарушению функции андрогенового рецептора вплоть до полной его инактивации.
Активация в клетках Лейдига биосинтеза тестостерона, необходимого для дифференцировки органов половой системы по мужскому типу происходит под действием транскрипционного активатора, кодируемого геном SFI (от англ. steroidogenic factor I), который локализован в длинном плече хромосомы 9 (9q33). Кроме того, предполагается, что ген SFI регулирует экспрессию гена DAXI, так как в промоторе последнего обнаружен распознающий SFI-сшг, т.е. по отношению к DAXI ген SF1 выступает в качестве «вышестоящего». В свою очередь DAX1 может быть супрессором в отношении гена SF1. Возможно, в процессе морфогенеза яичников ген DAXI предотвращает транскрипцию гена SOX9 через репрессию транскрипции гена SF1. Имеющиеся данные говорят о детерминирующей роли гена SF1 в дифференцировке эндокринных органов, регулирующее действие которых на морфогенез половой системы начинается после завершения ранних этапов дифференцировки гонад.
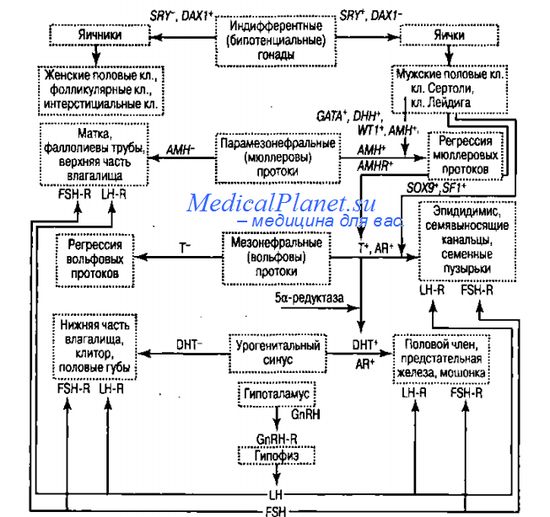
Кроме описанных выше гормонов, во вторичной детерминации пола у млекопитающих и, в частности, у человека, важную роль играют эстрогены и их рецепторы. Нормальная функция эстрогенов необходима для формирования репродуктивной системы как у женщин, так и у мужчин. Причем у последних эстрогены участвуют в созревании костной ткани и обеспечении определенных качественных показателей сперматозоидов. «Выключение» эстрогеновых рецепторов вызывает у модельных животных (мышей) гипоплазию матки и яичников в сочетании с развитием бесплодия.
Таким образом, в окончательном формировании наружных гениталий решающую роль играют андрогены — мужские половые гормоны, продуцирующиеся в надпочечниках и в яичках. Если их влияние отсутствует либо недостаточно, наружные половые органы формируются по женскому типу независимо от наличия или отсутствия эстрогенов.
Количество уже сейчас известных генов, вовлеченных в развитие и функционирование органов репродуктивной системы, чрезвычайно велико. Для яичка (как и для простаты) оно составляет более 1200, для яичника -более 500, для матки - более 1800 генов. Анализ функциональных взаимоотношений даже небольшой части наследственных детерминант, включенных в формирование пола у человека, позволяет получить представление о многоплановом их взаимодействии в данном процессе, который нельзя представить как цепь последовательных событий, подобную описанной выше у дрозофилы, По мнению ряда исследователей, процесс взаимодействия генетических регуляторов половой дифференцировки у человека, скорее всего, можно сравнить с сетью, где действие всех генов взаимосвязано и взаимообусловлено.
Рассматривая проблемы детерминации пола у человека, мы затронули лишь надводную часть «айсберга», каковым представляется чрезвычайно сложный процесс становления пола. Он может быть продемонстрирован схемой уровней дифференцировки пола, предложенной в 1990 г. Г.С. Васильченко.
Нарушение процесса формирования каждого из обозначенных уровней может стать у человека причиной отклонений в становлении половой идентичности.
- Читать далее "Зависимые от пола признаки. Отграниченные от пола признаки."
Оглавление темы "Пол и гены. Репликация генетического материала.":1. Пол. Дифференцировка пола. Основные виды детерминации пола.
2. Хромосомная детерминация пола. Гинандроморфизм.
3. Баланс полов. Балансовая теория Бриджерса.
4. Гены изменяющие пол. Механизмы детерминации пола у дрозофилы.
5. Пол у человека. Детерминация пола у человека. Роль Y хромосомы в детерминации пола.
6. Аутосомные гены в детерминации пола. Вторичная детерминация пола у человека.
7. Этапы вторичной детерминация пола. Стадии вторичной детерминации пола у человека.
8. Зависимые от пола признаки. Отграниченные от пола признаки.
9. Репликация генетических молекул. Полуконсервативная репликация ДНК и хромосом.
10. Типы репликации геномов. Инициация репликации генома.